

Epigenetic silencing and CRISPR-mediated reactivation of tight junction protein claudin10b (CLDN10B) in renal cancer
- PMID: 40524239
- PMCID: PMC12172364
- DOI: 10.1186/s13148-025-01911-2
Epigenetic silencing and CRISPR-mediated reactivation of tight junction protein claudin10b (CLDN10B) in renal cancer
Abstract
Background: The kidney's tubular system relies on cell polarity and tight junctions to maintain structure and function and disruptions contribute to diseases like cancer. Loss of tight junction proteins such as Claudins can actively contribute to tumorigenesis.
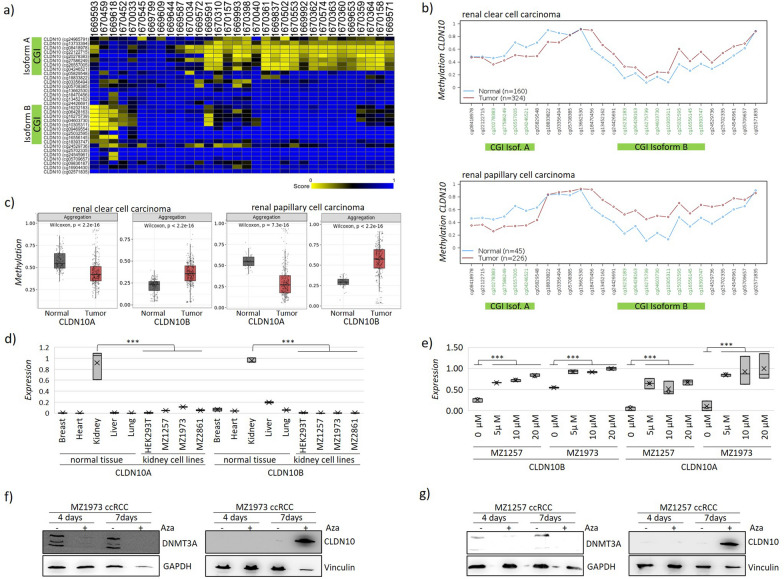
Results: We aimed to identify biomarkers for renal carcinoma, after kidney transplantation and conventional kidney tumors. We identified the epigenetic silencing of the Claudin 10 gene isoform B (CLDN10B) through DNA hypermethylation in renal cancers, including clear cell (ccRCC), papillary (pRCC) and post-transplantation renal carcinoma (PT-ccRCC). In contrast, CLDN10A was hypomethylated in ccRCC and pRCC. Differential methylation of the isoforms discriminates RCC from other malignancies. The epigenetic alteration of CLDN10B significantly correlated with reduced patient survival and advanced tumor staging. CLDN10B overexpression or induction significantly inhibited migration, cell cycle progression, and cellular growth. Using a CRISPR-based epigenetic editing tool reactivated CLDN10B to its endogenous level using VP160 and TET1 by promoter demethylation and significantly demonstrated its tumor-suppressive effects in 2D and 3D cell models.
Conclusion: Our findings suggest that CLDN10B acts as a tumor suppressor, and its epigenetic regulation may represent a therapeutic target for RCC. Ultimately, understanding CLDN10B's regulation and function could provide new insights into renal cancer treatment.
Keywords: CLDN10; CRISPR-Cas9; DNA (hyper)methylation; Epigenetic editing; Renal cell carcinoma (RCC); Tumor suppressor.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval: This study was funded by PTM samples from University Hospital Leipzig (J. Halbritter) with ethics approval (#213–19-ek). Competing interests: The authors declare no competing interests.
Figures






References
-
- Sung H, et al. Global cancer statistics 2020: globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49. - PubMed
-
- Siegel RL, et al. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17–48. - PubMed
-
- Siegel RL, et al. Cancer statistics, 2016. CA Cancer J Clin. 2016;66(1):7–30. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
