

Multi-step pathway engineering in probiotic Saccharomyces boulardii for abscisic acid production in the gut
- PMID: 40530243
- PMCID: PMC12173631
- DOI: 10.1016/j.mec.2025.e00263
Multi-step pathway engineering in probiotic Saccharomyces boulardii for abscisic acid production in the gut
Abstract
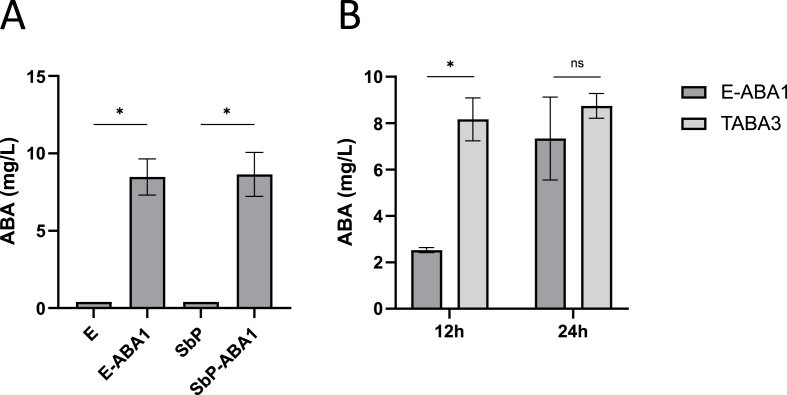
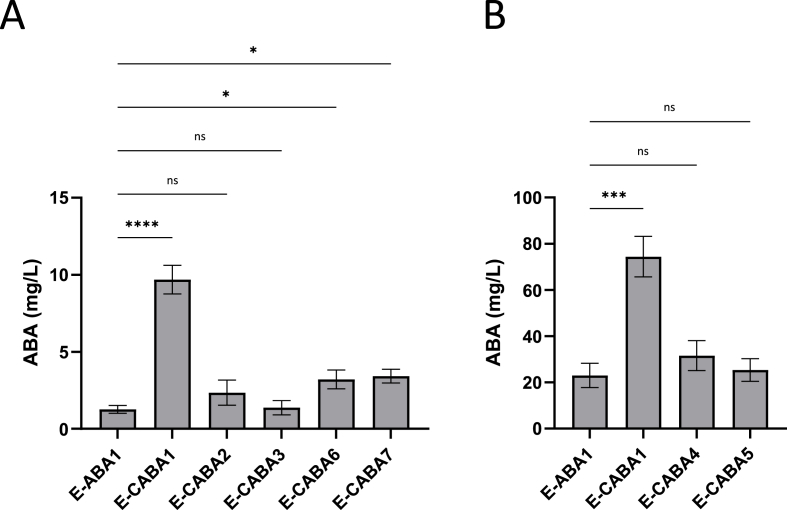
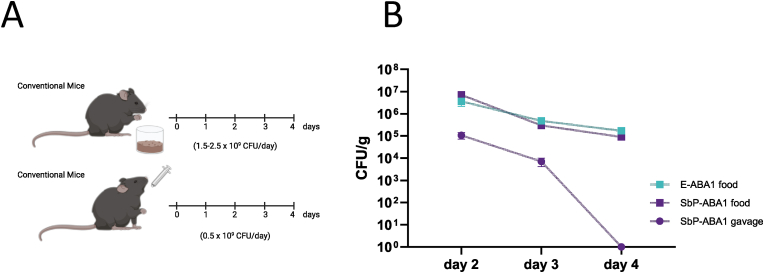
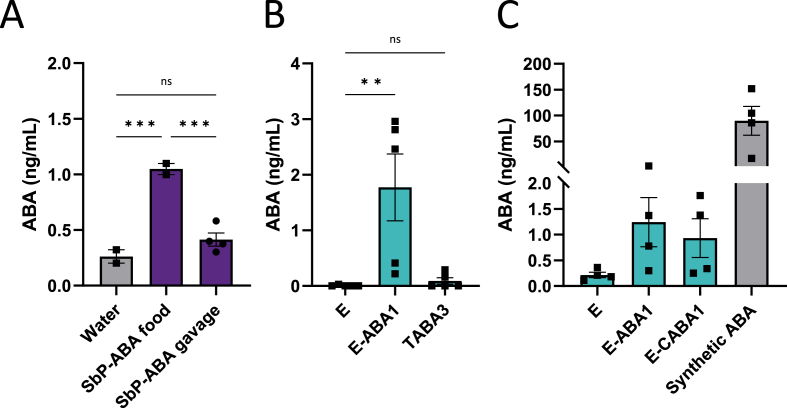
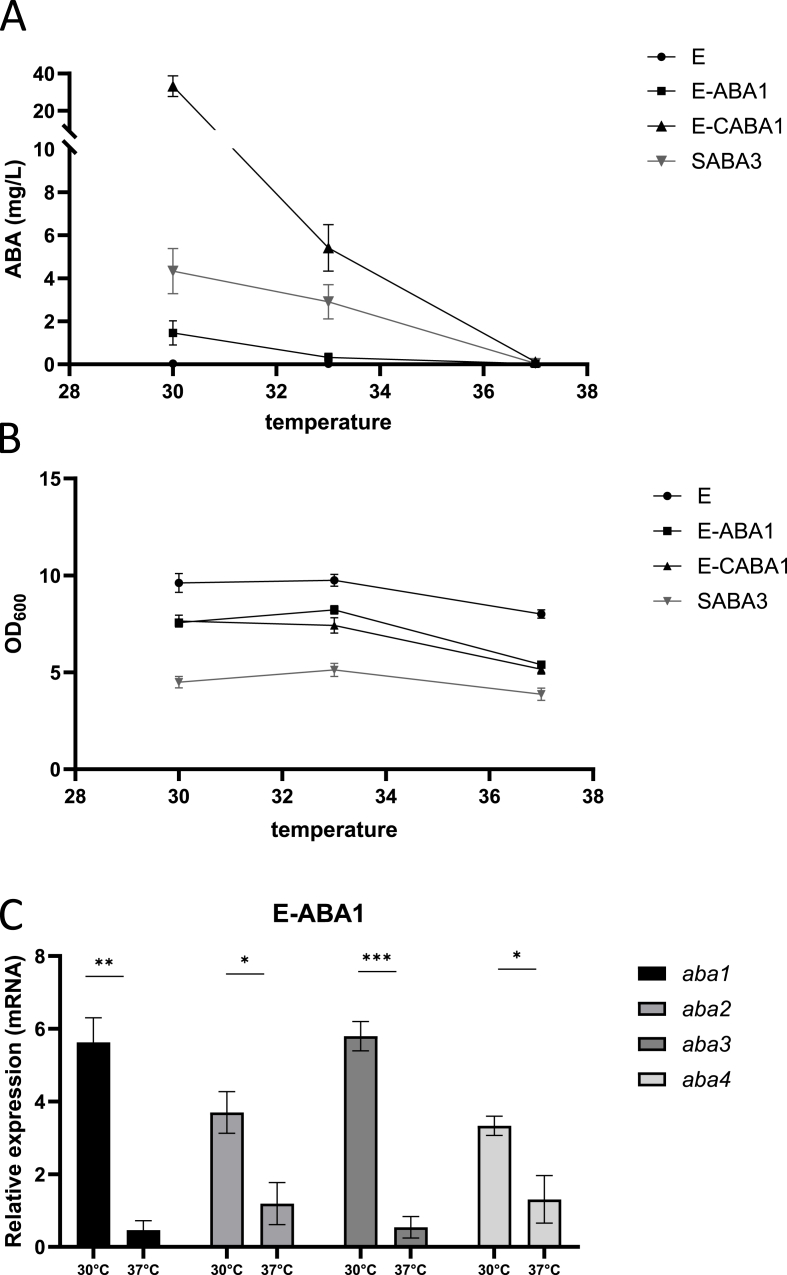
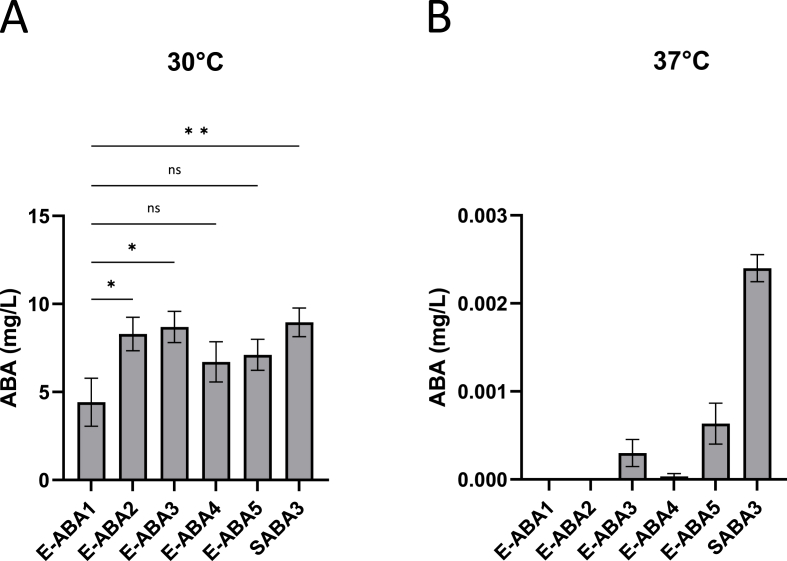
The plant hormone abscisic acid (ABA) has gained attention for its role in animals and humans, particularly due to its protective effects in various immune and inflammatory disorders. Given its high concentrations in fruits like figs, bilberries and apricots, ABA shows promise as a nutraceutical. However scalability, short half-life and cost limit the use of ABA-enriched fruit extracts and synthetic supplements. In this study, we propose an alternative ABA administration method to overcome these challenges. We genetically engineered a strain of the probiotic Saccharomyces boulardii to produce and deliver ABA directly to the gut of mice. Using the biosynthesis pathway from Botrytis cinerea, four genes (bcaba1-4) were integrated into S. boulardii, enabling ABA production at 30 °C, as previously described in Saccharomyces cerevisiae. Introducing an additional cytochrome P450 reductase gene resulted in a 7-fold increase in ABA titers, surpassing previous ABA-producing S. cerevisiae strains. Supplementation of the ABA-producing S. boulardii in the diet of mice (at a concentration of 5 × 108 CFU/g) led to effective gut colonization but resulted in low serum ABA levels (approximately 1.8 ng/mL). The absence of detectable serum ABA after administration of the ABA-producing probiotic through oral gavage, prompted further investigation to determine the underlying cause. The physiological body temperature (37 °C) was identified as a major bottleneck for ABA production. Modifications to enhance the mevalonate pathway flux improved ABA levels at 37 °C. However, additional modifications are needed to optimize ABA production before testing this probiotic in disease contexts in mice.
Keywords: Abscisic acid; In situ production; Metabolic engineering; Probiotic engineering; Saccharomyces boulardii.
© 2025 The Authors.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures

Similar articles
-
In Situ Biomanufacturing of Small Molecules in the Mammalian Gut by Probiotic Saccharomyces boulardii.ACS Synth Biol. 2021 May 21;10(5):1039-1052. doi: 10.1021/acssynbio.0c00562. Epub 2021 Apr 12. ACS Synth Biol. 2021. PMID: 33843197
-
Systematic Engineering for Efficient Uric Acid-Degrading Activity in Probiotic Yeast Saccharomyces boulardii.ACS Synth Biol. 2025 Jun 20;14(6):2030-2043. doi: 10.1021/acssynbio.4c00831. Epub 2025 May 8. ACS Synth Biol. 2025. PMID: 40340401 Free PMC article.
-
Genetic engineering of Saccharomyces boulardii: Tools, strategies and advances for enhanced probiotic and therapeutic applications.Biotechnol Adv. 2025 Jul 30;84:108663. doi: 10.1016/j.biotechadv.2025.108663. Online ahead of print. Biotechnol Adv. 2025. PMID: 40750061 Review.
-
Engineered Probiotic Saccharomyces boulardii Reduces Colitis-Associated Colorectal Cancer Burden in Mice.Dig Dis Sci. 2025 Jul;70(7):2348-2367. doi: 10.1007/s10620-025-09008-9. Epub 2025 Mar 29. Dig Dis Sci. 2025. PMID: 40156662 Free PMC article.
-
Molecular Genetics and Probiotic Mechanisms of Saccharomyces cerevisiae var. boulardii.Probiotics Antimicrob Proteins. 2025 Jul 24. doi: 10.1007/s12602-025-10634-y. Online ahead of print. Probiotics Antimicrob Proteins. 2025. PMID: 40705231 Review.
References
-
- Albenberg L., Esipova T.V., Judge C.P., Bittinger K., Chen J., Laughlin A., Grunberg S., Baldassano R.N., Lewis J.D., Li H., Thom S.R., Bushman F.D., Vinogradov S.A., Wu G.D. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology (New York, N. Y., 1943) 2014;147:1055–1063.e8. doi: 10.1053/j.gastro.2014.07.020. - DOI - PMC - PubMed
-
- Ameri P., Bruzzone S., Mannino E., Sociali G., Andraghetti G., Salis A., Ponta M.L., Briatore L., Adami G.F., Ferraiolo A., Venturini P.L., Maggi D., Cordera R., Murialdo G., Zocchi E. Impaired increase of plasma abscisic Acid in response to oral glucose load in type 2 diabetes and in gestational diabetes. PLoS One. 2015;10 doi: 10.1371/journal.pone.0115992. - DOI - PMC - PubMed
-
- Bakkes P.J., Riehm J.L., Sagadin T., Rühlmann A., Schubert P., Biemann S., Girhard M., Hutter M.C., Bernhardt R., Urlacher V.B. Engineering of versatile redox partner fusions that support monooxygenase activity of functionally diverse cytochrome P450s. Sci. Rep. 2017;7:9570. doi: 10.1038/s41598-017-10075-w. - DOI - PMC - PubMed
-
- Bruzzone S., Moreschi I., Usai C., Guida L., Damonte G., Salis A., Scarfì S., Millo E., De Flora A., Zocchi E. Abscisic acid is an endogenous cytokine in human granulocytes with cyclic ADP-ribose as second messenger. Proc. Natl. Acad. Sci. U. S. A. 2007;104:5759–5764. doi: 10.1073/pnas.0609379104. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
