

Structure-Based Design of PROTACS for the Degradation of Soluble Epoxide Hydrolase
- PMID: 40532036
- PMCID: PMC12257526
- DOI: 10.1021/acs.jmedchem.5c00552
Structure-Based Design of PROTACS for the Degradation of Soluble Epoxide Hydrolase
Abstract
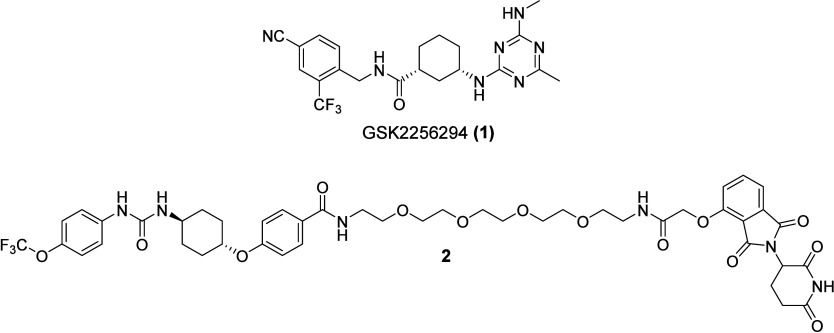
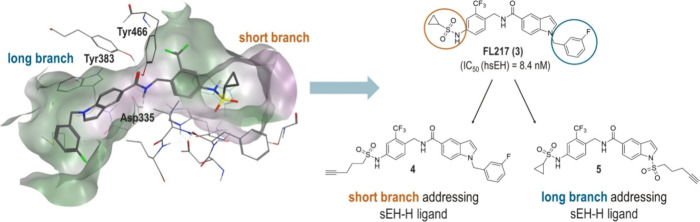
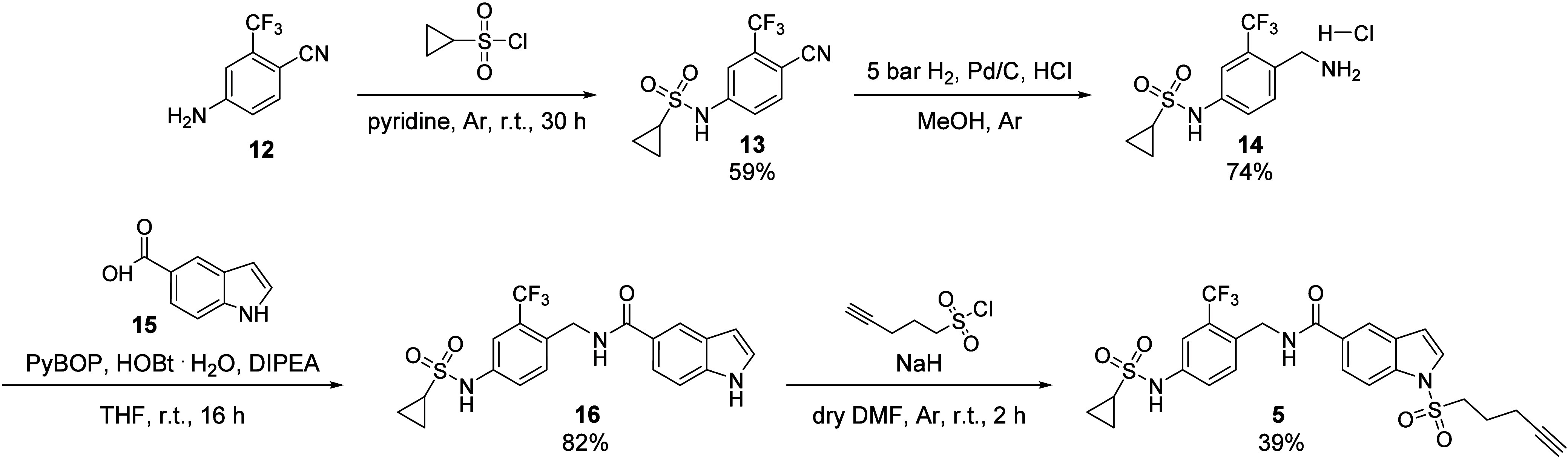
The bifunctional soluble epoxide hydrolase (sEH) represents a promising target for inflammation-related diseases. Although potent inhibitors targeting each domain are available, sEH-PROTACs offer the unique ability to simultaneously block both enzymatic functions, mimicking the sEH knockout phenotype, which has been associated with reducing inflammation, including neuroinflammation, and delaying the progression of Alzheimer's disease. Herein, we report the structure-based development of a potent sEH-PROTAC as a useful pharmacological tool. In order to facilitate a rapid testing of the PROTACs, a cell-based sEH degradation assay was developed utilizing HiBiT technology. We designed and synthesized 24 PROTACs. Furthermore, cocrystallization of sEH with two selected PROTACs allowed us to explore the binding mode and rationalize the most optimal linker length. After comprehensive biological and physicochemical characterization of this series, the most optimal PROTAC 23 was identified in primary human and murine cells, highlighting the potential of using 23 in disease-relevant cell and tissue models.
Figures










References
-
- Obeme-Nmom J. I., Udenigwe C. C.. Soluble epoxide hydrolase: an emerging target for nutraceuticals against inflammation and oxidative stress. Curr. Opin. Food Sci. 2024;57:101174. doi: 10.1016/j.cofs.2024.101174. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
