

Reminiscent of the pre-diatom? A hitherto undescribed scaly bolidophyte Lepidoparma frigida gen. et sp. nov. in a new order Lepidoparmales based on morphology, phylogeny, and ecology
- PMID: 40536273
- PMCID: PMC12351369
- DOI: 10.1111/jpy.70043
Reminiscent of the pre-diatom? A hitherto undescribed scaly bolidophyte Lepidoparma frigida gen. et sp. nov. in a new order Lepidoparmales based on morphology, phylogeny, and ecology
Abstract
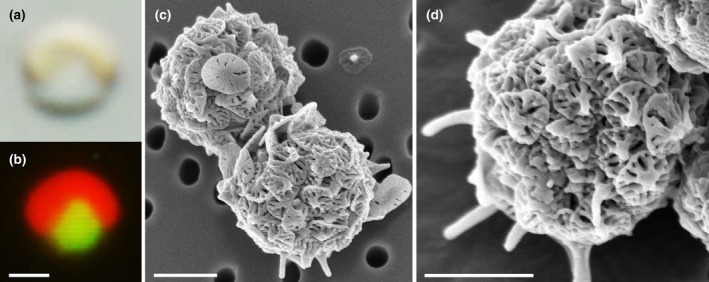
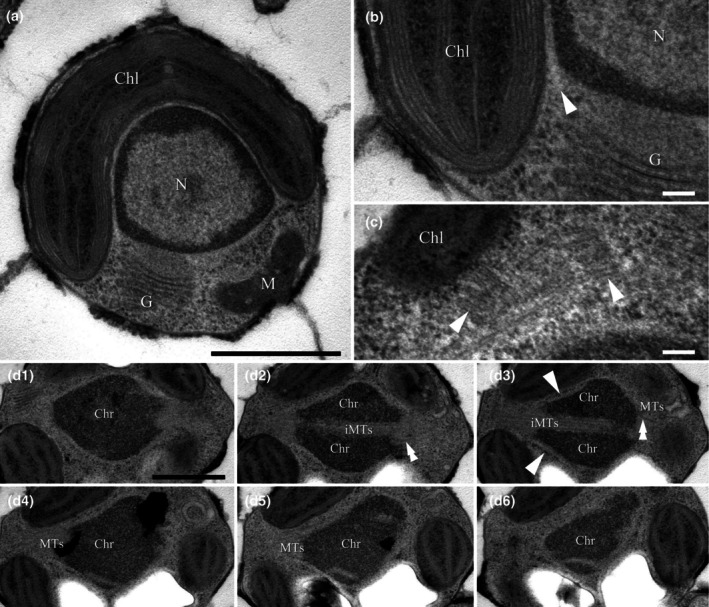
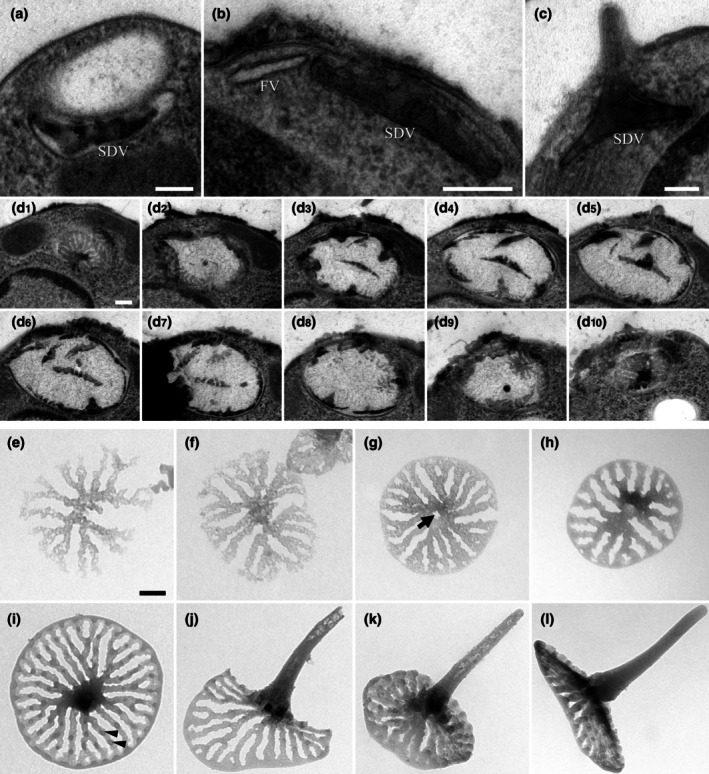
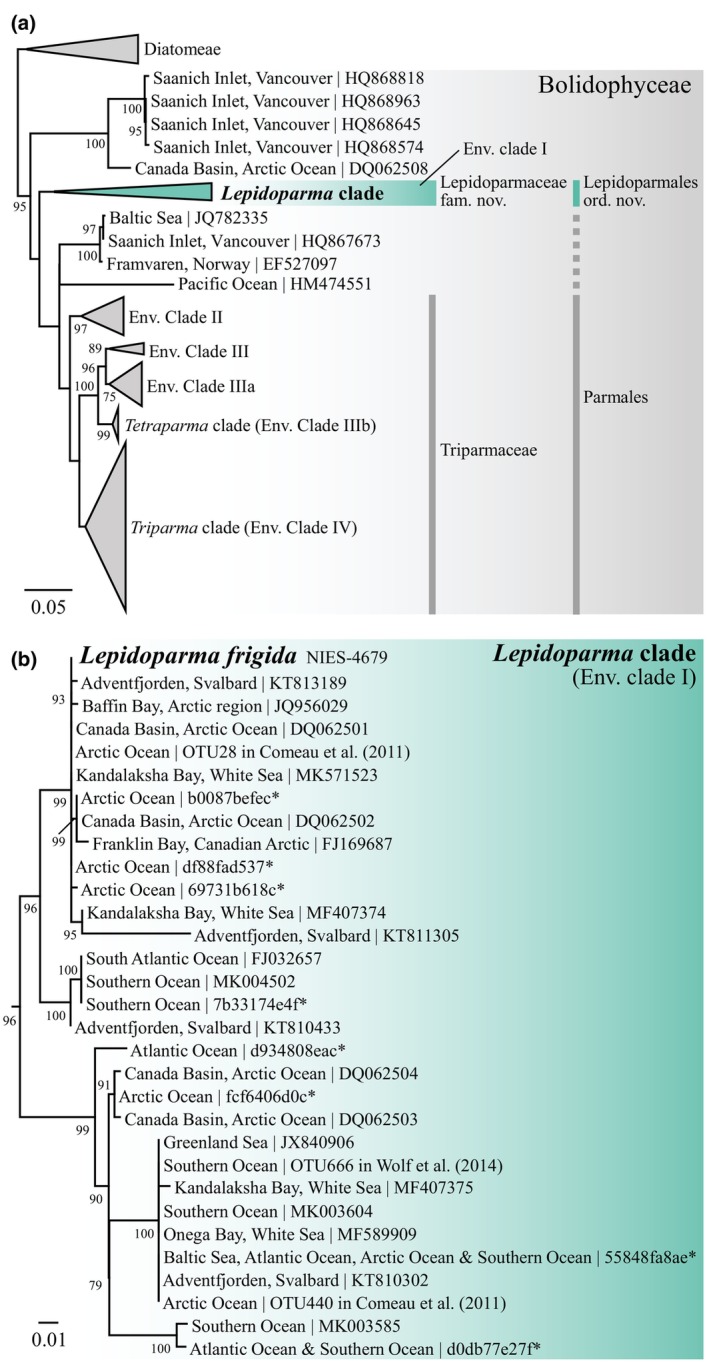
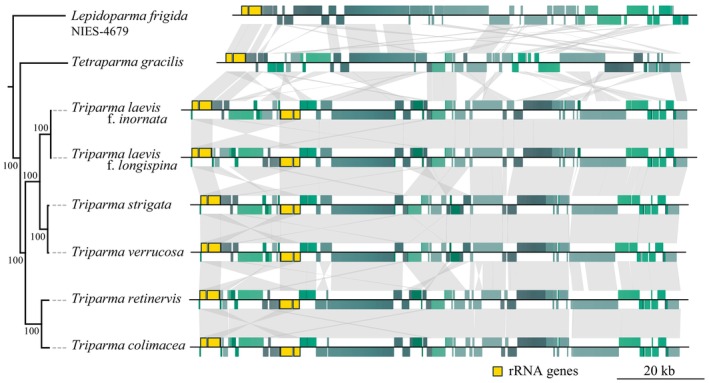
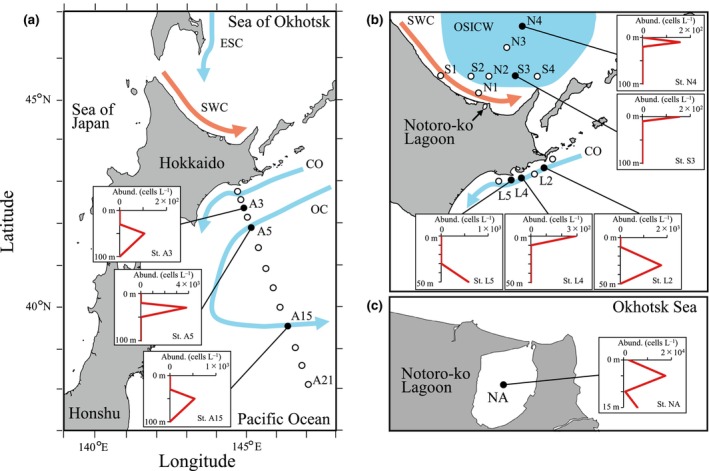
The class Bolidophyceae, which consists of small phytoplankton distributed worldwide, is the sister group of diatoms. This class has contained only one order, the Parmales, until now. In this study, we established a new order Lepidoparmales Kamakura & S.Sato ord. nov. and a new family Lepidoparmaceae Kamakura & S.Sato fam. nov., within the Bolidophyceae, and described Lepidoparma frigida Kamakura & S.Sato gen. et sp. nov. from the Sea of Okhotsk. We conducted a comprehensive comparison of L. frigida with diatoms and other parmaleans through analysis of its molecular phylogeny, morphology, ultrastructure, ontogeny, distribution, and lipid composition. The cell surface of L. frigida is covered with numerous siliceous scales with radial patterns similar to centric diatom valves; this character distinguishes it from other known parmaleans. Intriguingly, its appearance resembles the hypothesized "pre-diatom," which was proposed to be a precursor to primitive diatoms. The discovery and characterization of L. frigida will provide valuable insights into the evolutionary processes of both diatoms and bolidophytes and shed light on their common ancestor.
Keywords: biomineralization; marine phytoplankton; picoplankton; silica mineralization; taxonomy.
© 2025 The Author(s). Journal of Phycology published by Wiley Periodicals LLC on behalf of Phycological Society of America.
Figures
Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Determining dominant phytoplankton assemblages and their controlling factors in the winter sea-ice-covered southern sea of okhotsk and the spring open water through a multiple analytical approach.Mar Environ Res. 2025 Sep;210:107297. doi: 10.1016/j.marenvres.2025.107297. Epub 2025 Jun 14. Mar Environ Res. 2025. PMID: 40577886
-
Two novel Patescibacteria: Phycocordibacter aenigmaticus gen. nov. sp. nov. and Minusculum obligatum gen. nov. sp. nov., both associated with microalgae optimized for carbon dioxide sequestration from flue gas.mBio. 2025 Jul 9;16(7):e0123125. doi: 10.1128/mbio.01231-25. Epub 2025 Jun 12. mBio. 2025. PMID: 40503882 Free PMC article.
-
The medico-legal interpretation of diatom findings for the diagnosis of fatal drowning: a systematic review.Int J Legal Med. 2025 Mar;139(2):729-746. doi: 10.1007/s00414-024-03397-8. Epub 2025 Jan 14. Int J Legal Med. 2025. PMID: 39808189 Free PMC article.
-
[Volume and health outcomes: evidence from systematic reviews and from evaluation of Italian hospital data].Epidemiol Prev. 2013 Mar-Jun;37(2-3 Suppl 2):1-100. Epidemiol Prev. 2013. PMID: 23851286 Italian.
References
-
- Babenko, I. , Friedrich, B. M. , & Kröger, N. (2022). Structure and morphogenesis of the frustule. In Falciatore A. & Mock T. (Eds.), The molecular life of diatoms (pp. 287–312). Springer International Publishing. 10.1007/978-3-030-92499-7_11 - DOI
-
- Ban, H. , Sato, S. , Yoshikawa, S. , Yamada, K. , Nakamura, Y. , Ichinomiya, M. , Sato, N. , Blanc‐Mathieu, R. , Endo, H. , & Kuwata, A. (2023). Genome analysis of Parmales, the sister group of diatoms, reveals the evolutionary specialization of diatoms from phago‐mixotrophs to photoautotrophs. Communications Biology, 6(1), 697. 10.1038/s42003-023-05002-x - DOI - PMC - PubMed
-
- Bedoshvili, Y. D. , & Likhoshway, Y. V. (2019). Cellular mechanisms of diatom valve morphogenesis. In Seckbach J. & Gordon R. (Eds.), Diatoms: Fundamentals and applications (pp. 99–114). John Wiley & Sons, Ltd. 10.1002/9781119370741.ch5 - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
