

Facilitating Phloem-Mediated Iron Transport Can Improve the Adaptation of Rice Seedlings to Iron Deficiency Stress
- PMID: 40536616
- PMCID: PMC12179029
- DOI: 10.1186/s12284-025-00816-1
Facilitating Phloem-Mediated Iron Transport Can Improve the Adaptation of Rice Seedlings to Iron Deficiency Stress
Abstract
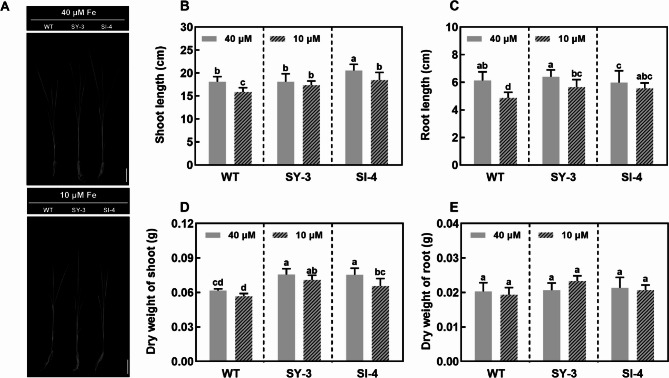
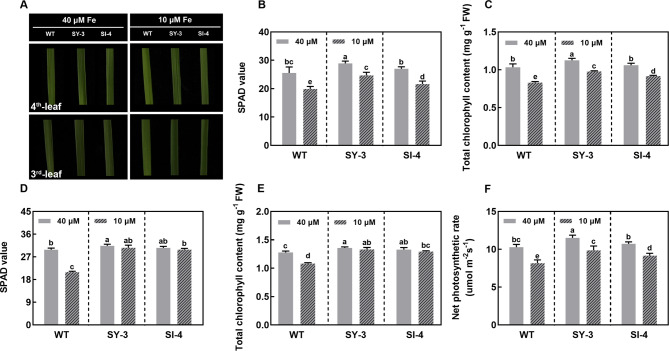
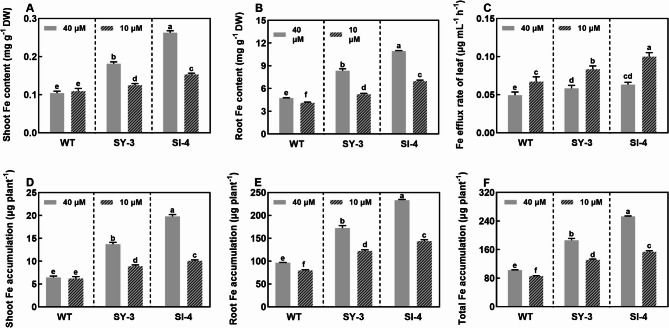
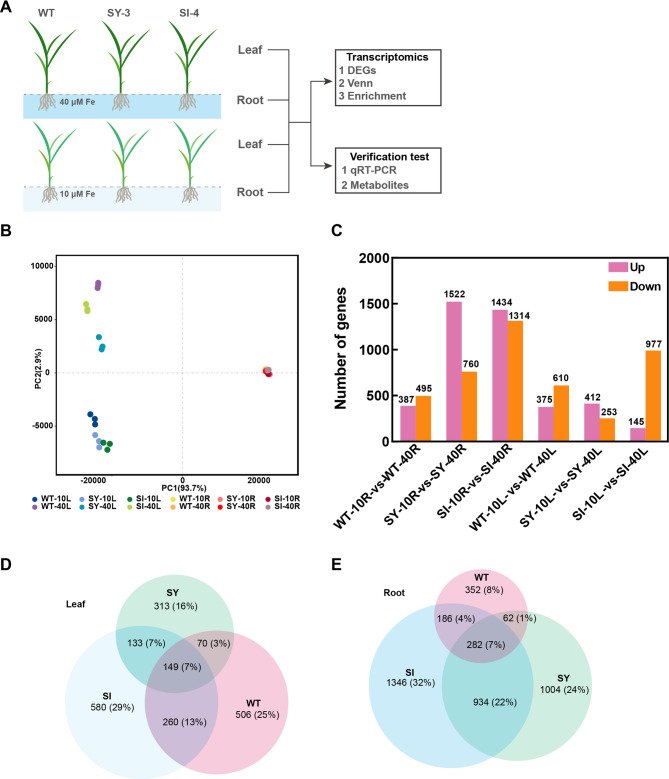
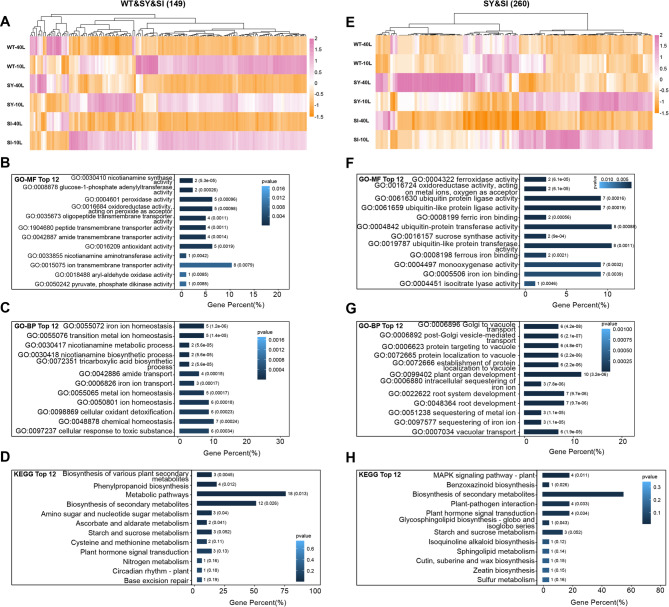
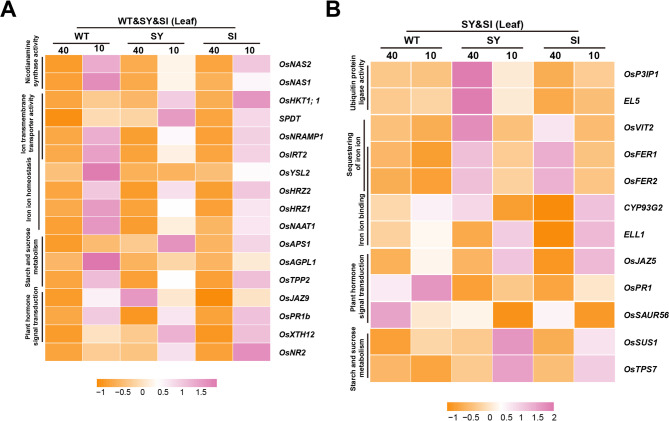
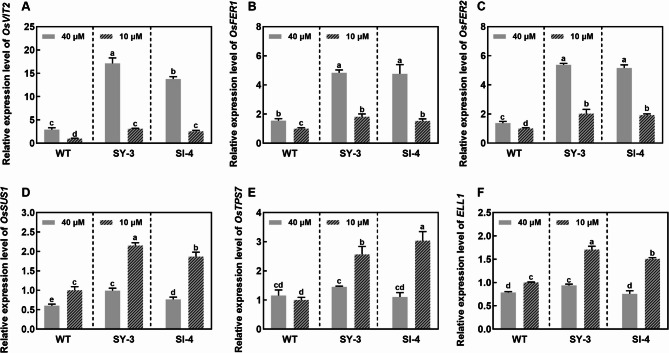
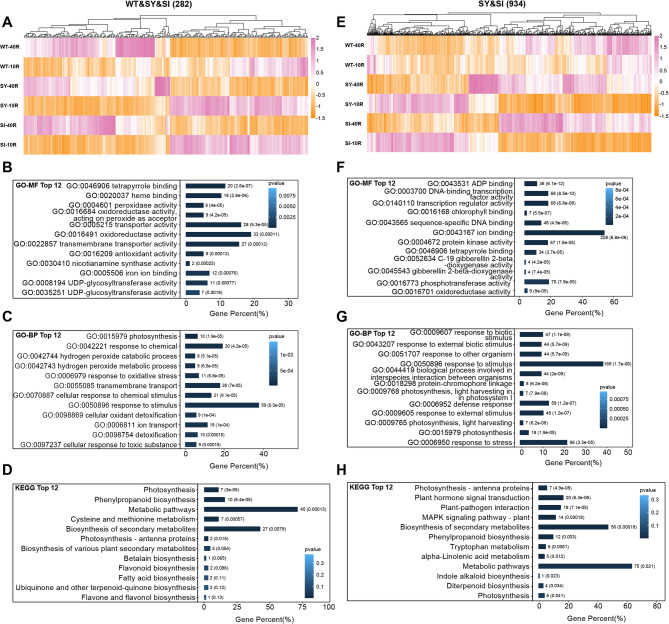
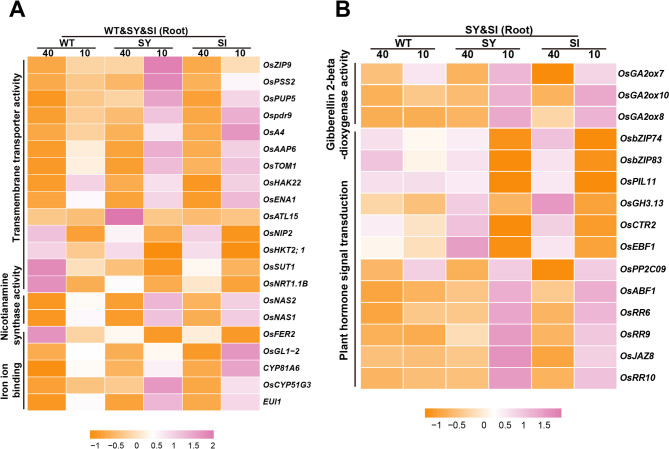
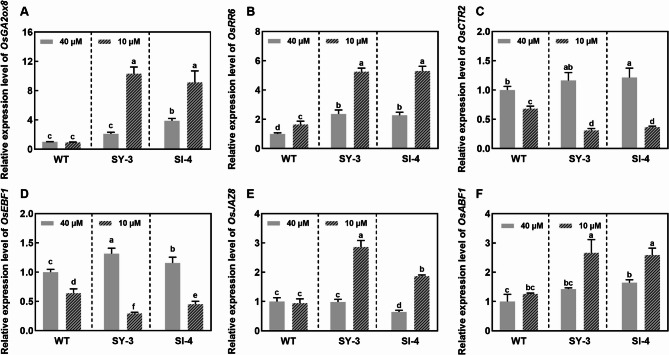
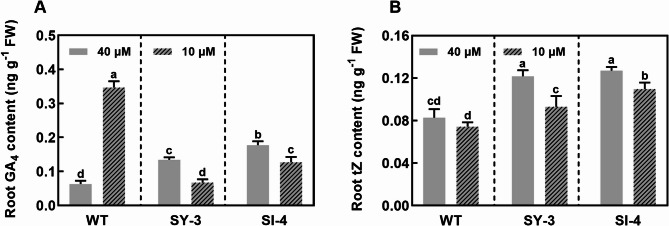
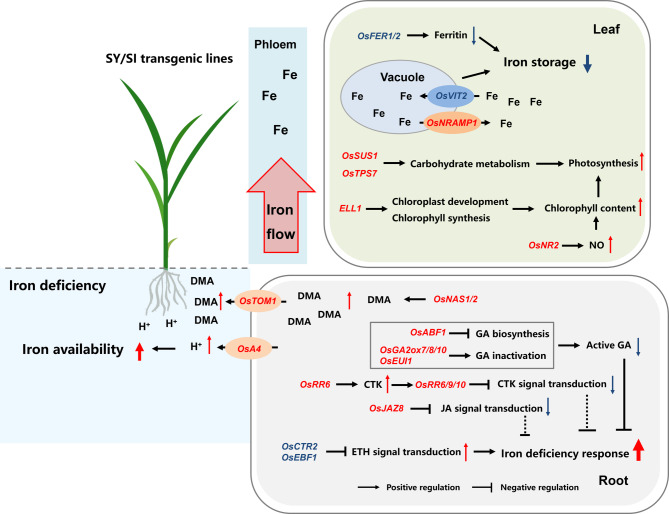
Iron (Fe) is essential for normal plant growth and development. In rice, Fe deficiency leads to stunted growth, leaf chlorosis, reduced photosynthetic capacity, and ultimately, yield loss. Most studies have focused on investigating the mechanisms of Fe deficiency responses in rice roots; however, the effects of shoot Fe redistribution on Fe deficiency response remain poorly understood. Phloem transport plays a vital role in distributing Fe to new tissues. To investigate the effects of enhanced phloem-mediated Fe transport on rice adaptability to iron deficiency, we subjected transgenic lines with higher phloem Fe efflux rates and wild-type (WT) plants to Fe-deficient conditions. The growth, leaf photosynthetic rate, and Fe content of transgenic and WT seedlings under different Fe concentrations were compared. The results showed that the transgenic lines exhibited elevated shoot length, root length, shoot dry weight, leaf chlorophyll content, and net photosynthetic rates under Fe-deficient conditions. Under both Fe-sufficient and Fe-deficient conditions, the transgenic lines had significantly higher Fe content, Fe accumulation, and phloem Fe efflux rates than the WT. RNA sequencing (RNA-seq) analysis revealed that enhanced Fe transport via phloem resulted in improved Fe availability through the sequestration of Fe ions and vacuolar transport pathways in the shoots. It also upregulated the EARLY LESION LEAF 1 (ELL1) expression and modulated the sucrose synthase activity, thereby promoting chlorophyll synthesis and leaf photosynthesis. Additionally, enhanced Fe transport influenced the gibberellin (GA) catabolism and plant hormone signal transduction in the roots, reducing the GA content and modulating the cytokinin (CTK), jasmonic acid (JA), and ethylene (ETH) signaling to induce Fe deficiency response and promote Fe uptake. These findings demonstrate that phloem-mediated Fe transport participated in Fe deficiency response, and enhancing this improved the adaptability of rice seedlings to low Fe conditions. In specific, rice seedlings with a high capacity for phloem-mediated Fe transport exhibited a strong iron uptake, translocation, and remobilization capacity, thereby maintaining normal growth and development and successfully adapting to the low-Fe environment.
Keywords: Iron Deficiency; Iron transport; Phloem; Plant hormone signal transduction; Rice (Oryza Sativa L.).
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Not applicable. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures
Similar articles
-
Impact of Iron Deficiency on the Arabidopsis thaliana Phloem Sap Proteome, a Key Role for bHLH121.Physiol Plant. 2025 May-Jun;177(3):e70336. doi: 10.1111/ppl.70336. Physiol Plant. 2025. PMID: 40536076 Free PMC article.
-
Integrated application of organic acids and earthworms improves rhizosphere microbiome, reduces health risks, and modulates proteomic and transcriptomic responses in rice (Oryza sativa L.) under aluminum stress.Ecotoxicol Environ Saf. 2025 Aug 7;303:118817. doi: 10.1016/j.ecoenv.2025.118817. Online ahead of print. Ecotoxicol Environ Saf. 2025. PMID: 40779847
-
Functional Analysis of Malus hallianaWRKY69 Transcription Factor (TF) Under Iron (Fe) Deficiency Stress.Curr Issues Mol Biol. 2025 Jul 21;47(7):576. doi: 10.3390/cimb47070576. Curr Issues Mol Biol. 2025. PMID: 40729045 Free PMC article.
-
Meta-Analysis of Iron Excess Stress in Rice: Genes and Mechanisms of Tolerance to Acidic Soil.Physiol Plant. 2025 Sep-Oct;177(5):e70473. doi: 10.1111/ppl.70473. Physiol Plant. 2025. PMID: 40873056 Free PMC article. Review.
-
A systematic review of single-cell RNA sequencing applications and insights in Oryza sativa response to environmental stresses.Mol Biol Rep. 2025 Jul 19;52(1):739. doi: 10.1007/s11033-025-10836-1. Mol Biol Rep. 2025. PMID: 40682700 Review.
References
-
- Aksoy E, Maqbool A, Tindas İ, Caliskan S (2017) Soybean: A new frontier in Understanding the iron deficiency tolerance mechanisms in plants. Plant Soil 418(1–2):37–44. 10.1007/s11104-016-3157-x
-
- Amir Hossain Md, Lee Y, Cho JI et al (2010) The bZIP transcription factor OsABF1 is an ABA responsive element binding factor that enhances abiotic stress signaling in rice. Plant Mol Biol 72(4–5):557–566. 10.1007/s11103-009-9592-9 - PubMed
-
- Bashir K, Inoue H, Nagasaka S et al (2006) Cloning and characterization of Deoxymugineic acid synthase genes from graminaceous plants. J Biol Chem 281:32395–32402. 10.1074/jbc.M604133200 - PubMed
-
- Briat JF, Duc C, Ravet K, Gaymard F (2010) Ferritins and iron storage in plants. Biochimica et biophysica acta (BBA). - Gen Subj 1800:806–814. 10.1016/j.bbagen.2009.12.003 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
