

Iron deficiency causes aspartate-sensitive dysfunction in CD8+ T cells
- PMID: 40541943
- PMCID: PMC12181297
- DOI: 10.1038/s41467-025-60204-7
Iron deficiency causes aspartate-sensitive dysfunction in CD8+ T cells
Abstract
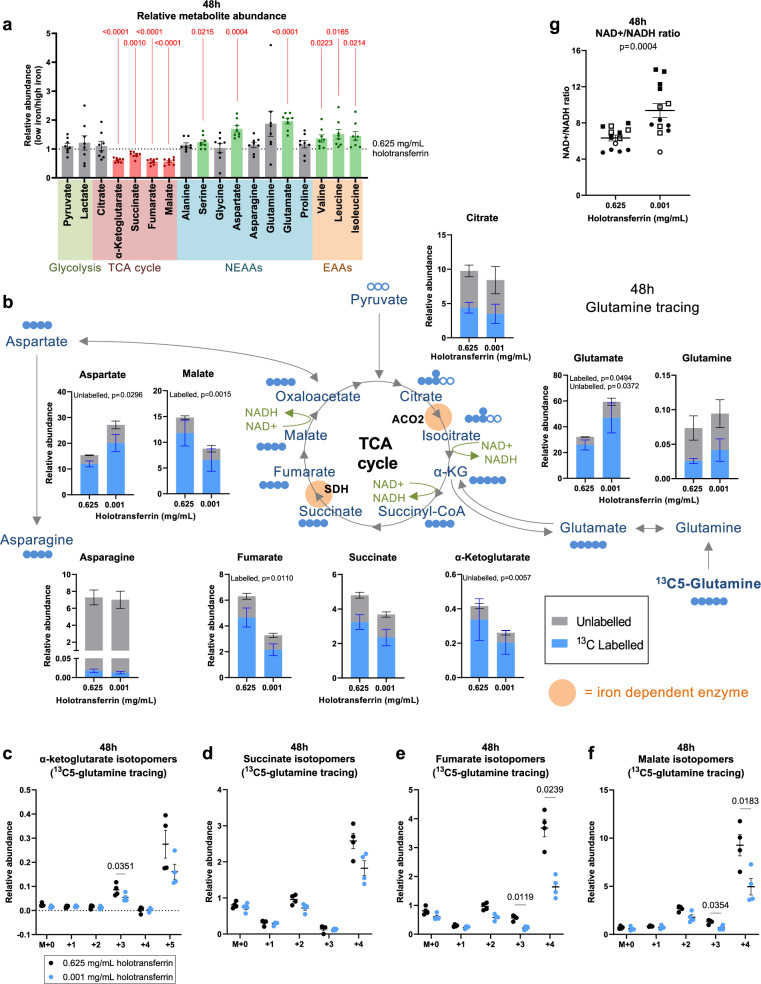
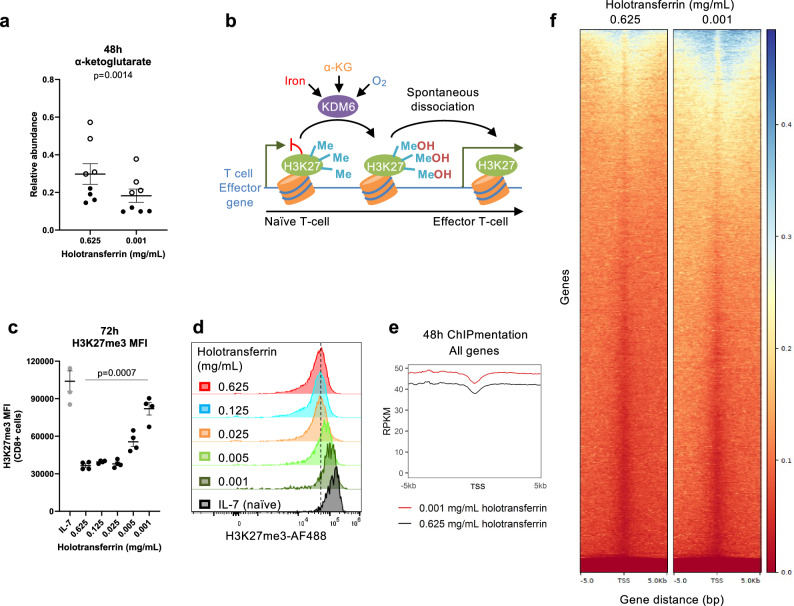
Iron is an irreplaceable co-factor for metabolism. Iron deficiency affects >1 billion people and decreased iron availability impairs immunity. Nevertheless, how iron deprivation impacts immune cell function remains poorly characterised. We interrogate how physiologically low iron availability affects CD8+ T cell metabolism and function, using multi-omic and metabolic labelling approaches. Iron limitation does not substantially alter initial post-activation increases in cell size and CD25 upregulation. However, low iron profoundly stalls proliferation (without influencing cell viability), alters histone methylation status, gene expression, and disrupts mitochondrial membrane potential. Glucose and glutamine metabolism in the TCA cycle is limited and partially reverses to a reductive trajectory. Previous studies identified mitochondria-derived aspartate as crucial for proliferation of transformed cells. Despite aberrant TCA cycling, aspartate is increased in stalled iron deficient CD8+ T cells but is not utilised for nucleotide synthesis, likely due to trapping within depolarised mitochondria. Exogenous aspartate markedly rescues expansion and some functions of severely iron-deficient CD8+ T cells. Overall, iron scarcity creates a mitochondrial-located metabolic bottleneck, which is bypassed by supplying inhibited biochemical processes with aspartate. These findings reveal molecular consequences of iron deficiency for CD8+ T cell function, providing mechanistic insight into the basis for immune impairment during iron deficiency.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: T.A.M. is a paid consultant for and shareholder in Dark Blue Therapeutics Ltd. D.A.T. undertakes paid consultancy work for Sitryx Ltd. The remaining authors declare no competing interests.
Figures




References
-
- Pasricha, S. R., Tye-Din, J., Muckenthaler, M. U. & Swinkels, D. W. Iron deficiency. Lancet397, 233–248 (2021). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
