

Acyl chains stabilize the acylated domain and determine the receptor-mediated interaction of the Bordetella adenylate cyclase toxin with cell membrane
- PMID: 40543585
- PMCID: PMC12332401
- DOI: 10.1016/j.jbc.2025.110392
Acyl chains stabilize the acylated domain and determine the receptor-mediated interaction of the Bordetella adenylate cyclase toxin with cell membrane
Abstract
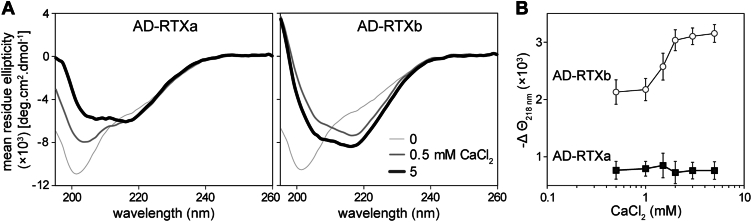
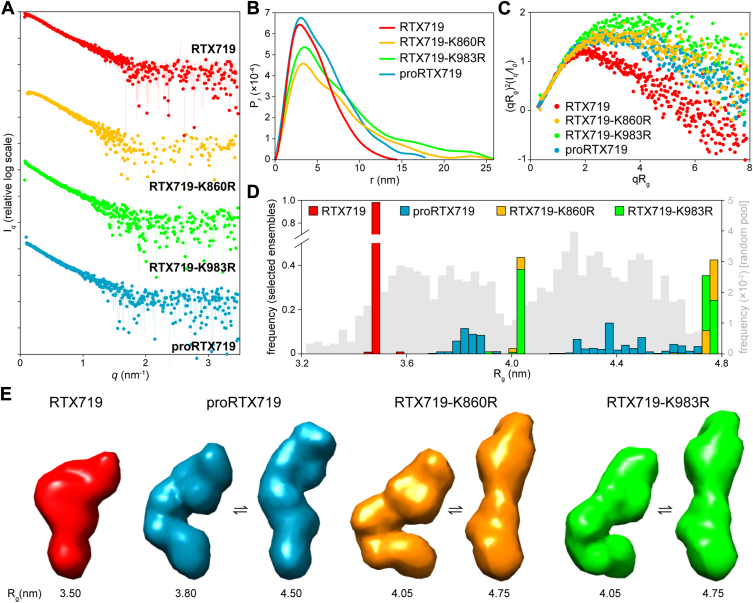
Acylated domains (ADs), like that of the Bordetella pertussis adenylate cyclase toxin (CyaA), are structures found in all pore-forming toxins from the family of Repeat-in-ToXin (RTX) proteins. These AD segments are fatty-acylated on ε-amino groups of conserved lysine residues, such as the K860 and K983 residues of CyaA. The ε-amide-linked acyl chains are essential for toxin activity and promote irreversible membrane insertion of the CyaA molecule, thus enabling the toxin to translocate its N-terminal adenyl cyclase enzyme domain into the host cell cytoplasm. In parallel, the membrane-inserted CyaA molecules can oligomerize into cation-selective pores in the plasma membrane. Here, we show that the attached acyl chains are not only crucial for membrane insertion of the toxin but also play an important role in CyaA folding. We demonstrate that assembly of the noncanonical β-roll structure in the C-terminal segment of the AD of CyaA is cooperatively directed by the Ca2+-driven folding of the adjacent RTX domain. In contrast, the N-terminal AD segment consists of an α-helical structure that folds independently of Ca2+ ion binding and may form one or two acyl binding site(s) accommodating the acyl chains protruding from the C-terminal AD segment. This acyl-mediated interaction between the N- and C-terminal segments promotes local structural rearrangements within the AD that significantly enhances the stability of the toxin molecule. These findings highlight the critical role of the acyl modification in membrane interaction capacity and structural stability of the CyaA toxin.
Keywords: Bordetella pertussis; RTX toxin; acylation; adenylate cyclase toxin; protein folding.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures




References
-
- El-Azami-El-Idrissi M., Bauche C., Loucka J., Osicka R., Sebo P., Ladant D., Leclerc C. Interaction of Bordetella pertussis adenylate cyclase with CD11b/CD18: role of toxin acylation and identification of the main integrin interaction domain. J. Biol. Chem. 2003;278:38514–38521. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
