

Gasdermin E in glioblastoma -pyroptosis resistance and tumor-promoting functions
- PMID: 40544161
- PMCID: PMC12182582
- DOI: 10.1038/s41420-025-02572-z
Gasdermin E in glioblastoma -pyroptosis resistance and tumor-promoting functions
Abstract
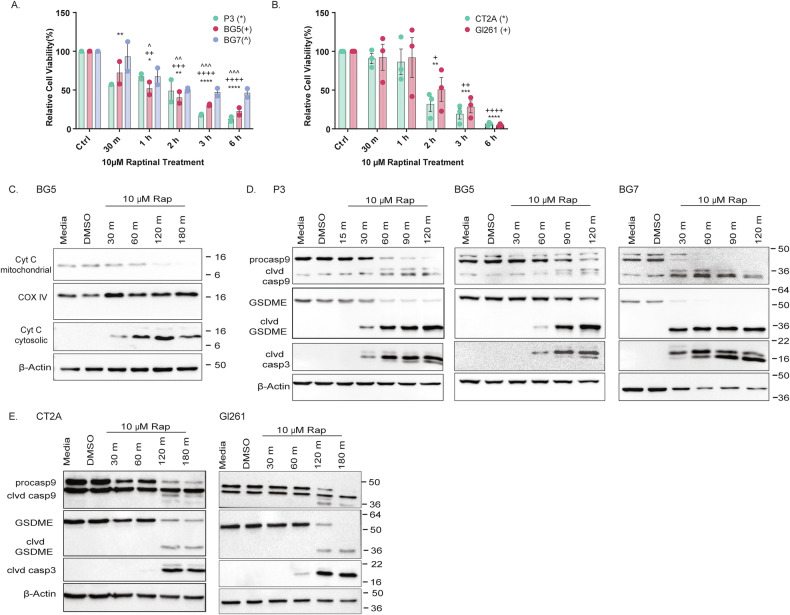
Treatment of glioblastoma (GB), the most common and most aggressive malignant brain tumor, has made little progress over the past two decades. Despite extensive research on apoptosis and autophagy, necrotic cell death mechanisms like pyroptosis, which have the potential to stimulate anti-tumor immune responses, remain largely underexplored in GB. Here, we investigated whether Gasdermin E (GSDME)-mediated pyroptosis can be induced in GB by employing the drug raptinal, an inducer of cytochrome c release. Using human patient-derived and mouse GB cell lines, we showed that raptinal promotes GSMDE cleavage. However, although a strong pyroptotic response was observed in mouse cell lines, it was weak in human cell lines. This resistance was partially reversed by the calcium chelator BAPTA-AM, indicating that membrane repair mechanisms may counteract the pyroptotic response. Gsdme knockout (KO) in mouse GB cells unexpectedly prolonged the survival of immunocompetent mice, demonstrating a tumor-promoting role of GSDME independent of its pyroptotic function. Analysis of the immune microenvironment revealed that Gsdme KO promoted infiltration of T cells, which was confirmed by spatial transcriptomic analysis of GB patient samples. In addition, Gsdme/GSMDE KO reduced the invasive capacity of mouse/human GB cells. In conclusion, active membrane repair mechanisms may impair the pyroptotic efficacy in GB. GSDME has a tumor-promoting role in GB by suppressing T cell infiltration and increasing tumor cell invasion.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests. Ethics statement: The study was conducted in accordance with the Declaration of Helsinki and approved by the Regional Ethical Committee (REK 151825, 10.09.2020). Informed consent was obtained from all subjects involved in the study. The Regional Ethics Committee approved the animal study protocol. Animals were treated in accordance with the Norwegian Animal Act.
Figures




References
-
- Ravi VM, Will P, Kueckelhaus J, Sun N, Joseph K, Salie H, et al. Spatially resolved multi-omics deciphers bidirectional tumor-host interdependence in glioblastoma. Cancer Cell. 2022;40:639–55.e13. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
