

Optogenetic perturbation of lipid droplet localization affects lipid metabolism and development in Drosophila
- PMID: 40545239
- PMCID: PMC12302289
- DOI: 10.1016/j.jlr.2025.100848
Optogenetic perturbation of lipid droplet localization affects lipid metabolism and development in Drosophila
Abstract
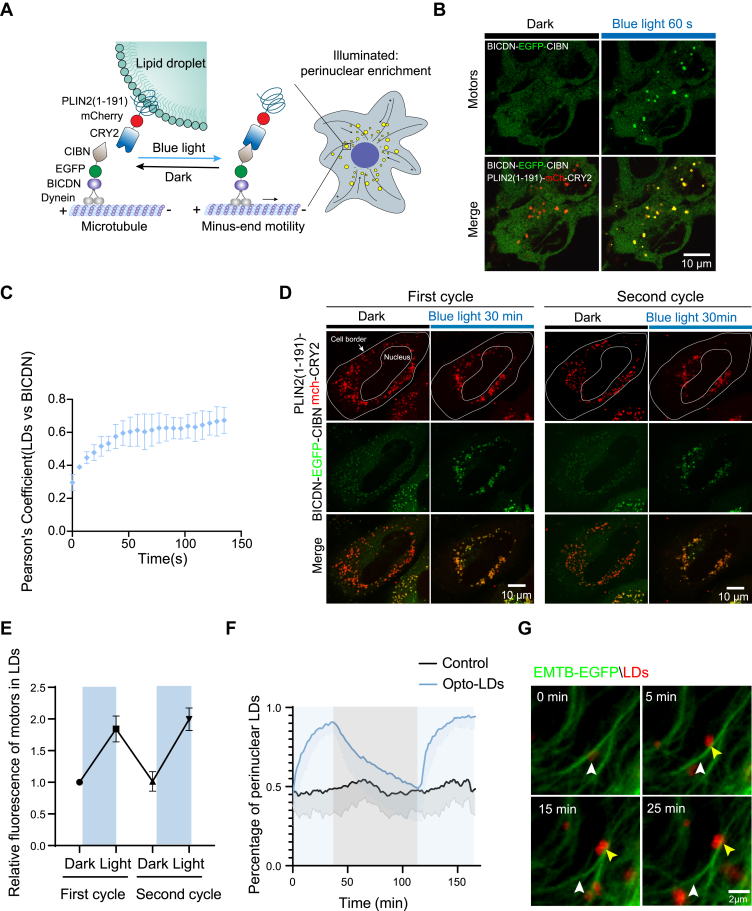
Lipid droplets (LDs) are dynamic organelles crucial for lipid storage and homeostasis. Despite extensive documentation of their importance, the causal relationship between LD localization and function in health and disease remains inadequately understood. Here, we developed optogenetics-based tools, termed "Opto-LDs," which facilitate the interaction between LDs and motor proteins in a light-dependent manner, enabling precise control of LD localization within cells. Utilizing these optogenetic modules, we demonstrated that light-induced relocation of LDs to the periphery of hepatocytes results in elevated very-low-density lipoprotein (VLDL) secretion, recapturing the beneficial effect of insulin in vitro. Furthermore, our studies in transgenic Drosophila revealed that proper LD localization is critical for embryonic development, with mistargeting of LDs significantly affecting egg hatching success. In summary, our work underscores the great importance of LD localization in lipid metabolism and development, and our developed tools offer valuable insights into the functions of LDs in health and disease.
Keywords: Drosophila; development; lipid droplet; metabolism; oogenesis; optogenetics.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures



References
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
