

Genetically distinct hantaviruses in two bat species in Panamá
- PMID: 40546951
- PMCID: PMC12178796
- DOI: 10.1016/j.isci.2025.112749
Genetically distinct hantaviruses in two bat species in Panamá
Abstract
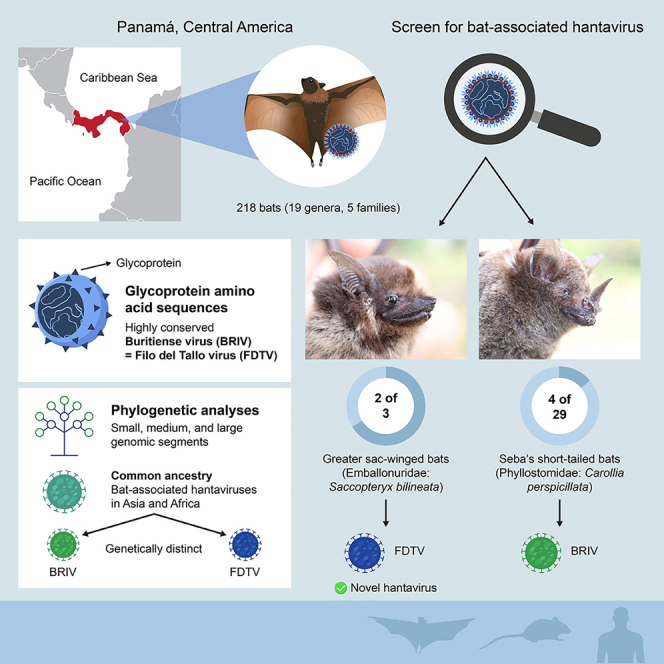
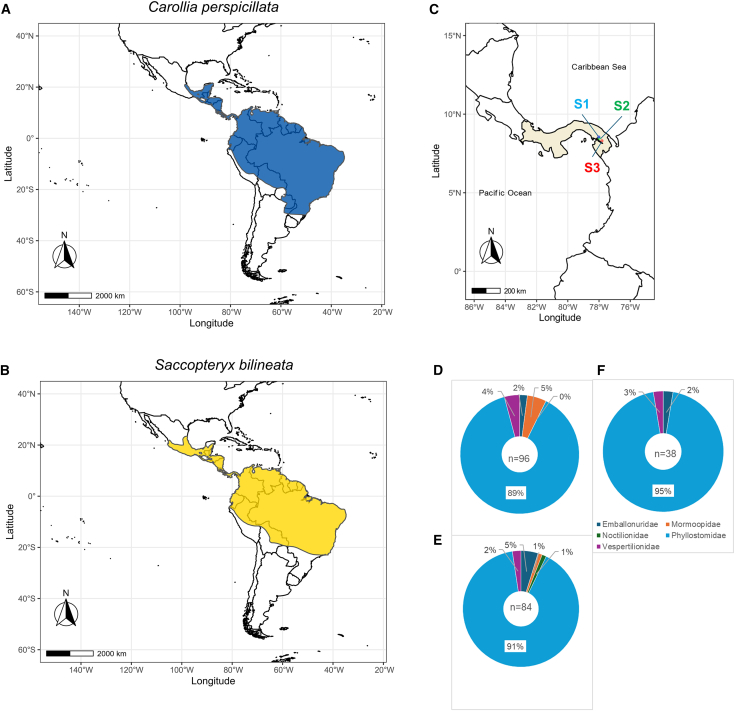
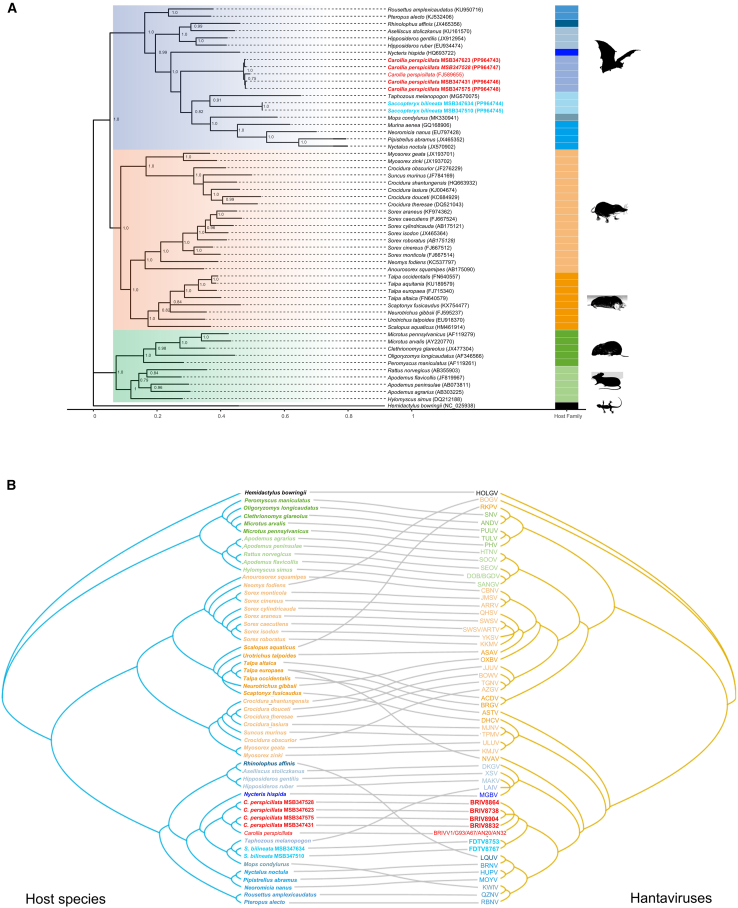
Recent discoveries of hantaviruses in bats in Europe, Asia, and Africa have prompted expanded explorations of their host diversity and geographic distribution. In screening lung tissue of 218 bats from Panamá, representing 19 genera and five families, we detected hantavirus RNA in two of three greater sac-winged bats (Emballonuridae: Saccopteryx bilineata) and in four of 29 Seba's short-tailed bats (Phyllostomidae: Carollia perspicillata). Phylogenetic analyses of the small, medium, and large genomic segments revealed a newfound hantavirus in S. bilineata, tentatively designated Filo del Tallo virus, and the recently reported Buritiense virus in C. perspicillata. Both share common ancestry with bat-associated hantaviruses in Asia and Africa. These results indicate that bats serve as hosts of hantaviruses in the Americas. Given opportunities for close contact between humans and bats, studies on the genetic diversity, pathogenicity, and circulation dynamics of hantaviruses are urgently needed.
Keywords: Microbiology; Virology; Zoology.
© 2025 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
A newly bat-borne hantavirus detected in Seba's short-tailed bats (Carollia perspicillata) in the Brazilian Atlantic Rainforest.Mem Inst Oswaldo Cruz. 2024 Dec 16;119:e240132. doi: 10.1590/0074-02760240132. eCollection 2024. Mem Inst Oswaldo Cruz. 2024. PMID: 39699489 Free PMC article.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
-
Major bat-borne zoonotic viral epidemics in Asia and Africa: A systematic review and meta-analysis.Vet Med Sci. 2022 Jul;8(4):1787-1801. doi: 10.1002/vms3.835. Epub 2022 May 10. Vet Med Sci. 2022. PMID: 35537080 Free PMC article.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
-
Factors that influence the provision of intrapartum and postnatal care by skilled birth attendants in low- and middle-income countries: a qualitative evidence synthesis.Cochrane Database Syst Rev. 2017 Nov 17;11(11):CD011558. doi: 10.1002/14651858.CD011558.pub2. Cochrane Database Syst Rev. 2017. PMID: 29148566 Free PMC article.
References
-
- Lee H.W., Lee P.W., Lähdevirta J., Brummer-Korventkontio M. Aetiological relation between Korean haemorrhagic fever and nephropathia epidemica. Lancet. 1979;1:186–187. - PubMed
-
- Yanagihara R., Gajdusek D.C. In: CRC Handbook of Viral and Rickettsial Hemorrhagic Fever. Gear J.H.S., editor. CRC Press; 1988. Hemorrhagic fever with renal syndrome: a historical perspective and review of recent advances; pp. 155–188.
-
- Nichol S.T., Spiropoulou C.F., Morzunov S., Rollin P.E., Ksiazek T.G., Feldmann H., Sanchez A., Childs J., Zaki S., Peters C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Sci. Technol. Humanit. 1993;262:914–917. - PubMed
-
- Yanagihara R., Gu S.H., Song J.-W. In: Global Virology I - Identifying and Investigating Viral Diseases. Shapshak P., Sinnott J.T., Somboonwit C., Kuhn J.H., editors. Springer; 2015. Expanded host diversity and global distribution of Hantaviruses: implications for identifying and investigating previously unrecognized Hantaviral diseases; pp. 161–198. - DOI
LinkOut - more resources
Full Text Sources
