

Evolutionary dynamics of heparan sulfate utilization by SARS-CoV-2
- PMID: 40548749
- PMCID: PMC12345153
- DOI: 10.1128/mbio.01303-25
Evolutionary dynamics of heparan sulfate utilization by SARS-CoV-2
Abstract
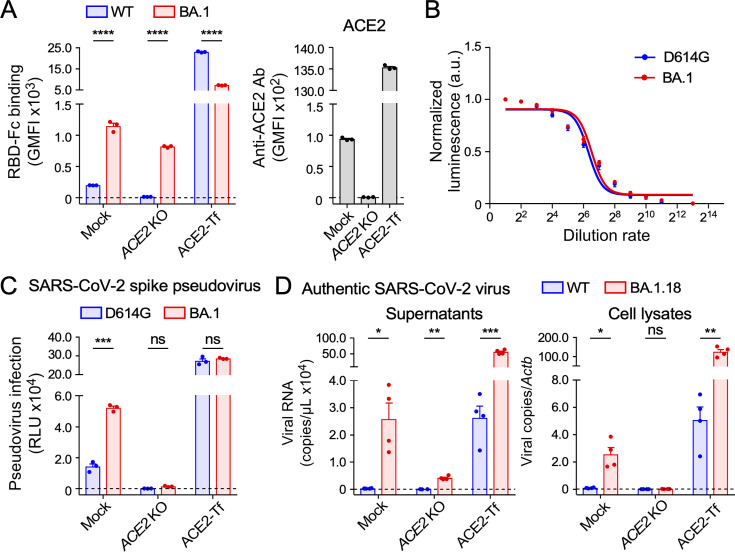
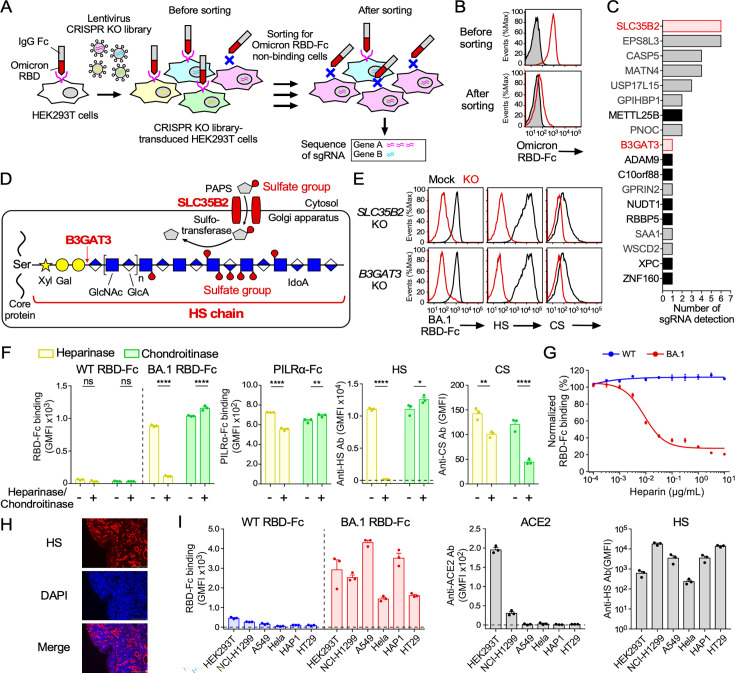
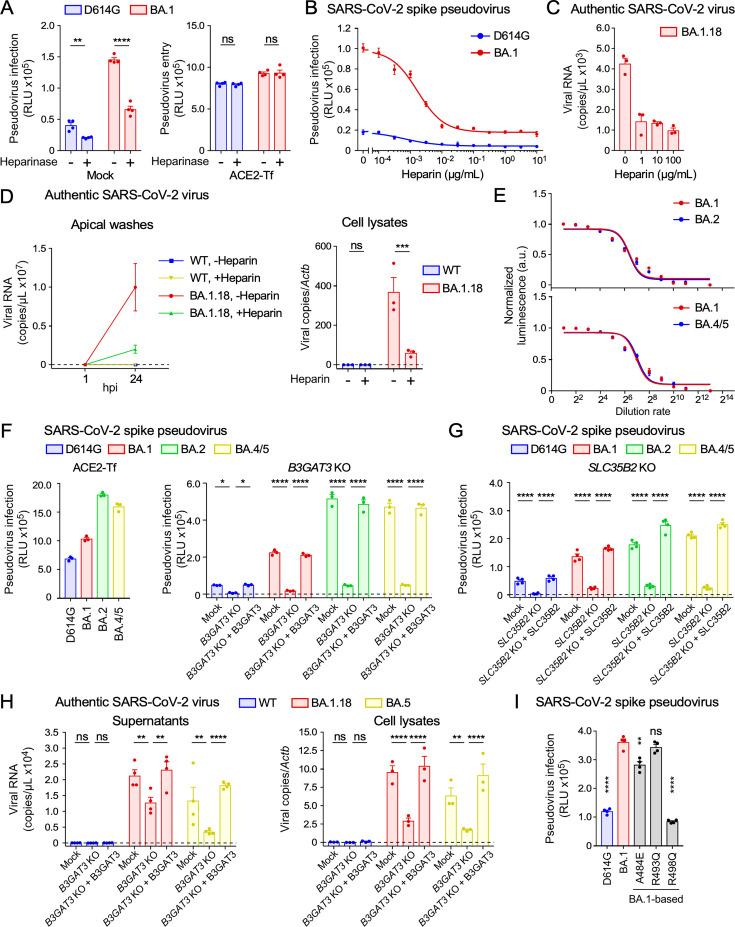
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Omicron variants have acquired enhanced infectivity compared to earlier variants. To elucidate the underlying molecular mechanisms, we conducted CRISPR library screening to identify cell surface molecules that interact with the Omicron spike protein. Our findings revealed a significantly higher affinity between the Omicron spike and cell surface heparan sulfate compared to the wild-type spike. This increased binding affinity enables Omicron variants to infect cells expressing low levels of ACE2, which are minimally infected by the wild-type virus. Mutational analysis of heparan sulfate binding sites on the Omicron spike protein, coupled with electrostatic potential mapping, suggested that the accumulation of positively charged mutations has contributed to the enhanced heparan sulfate binding. Comparative analysis of heparan sulfate binding among Omicron subvariants-including BA.1, BA.2, BA.4, BA.5, XBB.1, and BA.2.86-revealed that most are likely to bind efficiently to heparan sulfate, but potential heparan sulfate binding sites of the spike protein have shifted from the early Omicron variants to more recent ones. Furthermore, we discovered that cell surface heparan sulfate proteoglycans are cleaved by TMPRSS2, a protease essential for wild-type SARS-CoV-2 infection. These findings suggest that SARS-CoV-2 is evolving to enhance its infectivity by optimizing its interaction with cell surface heparan sulfate.IMPORTANCEThe Omicron variant has evolved to become highly infectious by acquiring numerous mutations. Understanding the impact of these mutations can provide valuable insights into the drivers of viral evolution and aid in the development of improved viral surveillance and vaccines. Our study demonstrates that the Omicron variants contain mutations that enhance their ability to bind to heparan sulfate. Highly infectious human viruses often utilize heparan sulfate for infection, suggesting that heparan sulfate likely plays a crucial role in viral adaptation to human hosts. Furthermore, we found that cell surface heparan sulfate proteoglycans are sensitive to TMPRSS2, while most other cell surface proteins are resistant to TMPRSS2. Given that TMPRSS2 is known to enhance the infectivity of earlier severe acute respiratory syndrome coronavirus 2 variants but cleaves heparan sulfate proteoglycans, it is probable that the high heparan sulfate binding acquired by the Omicron variant contributes to its decreased infectivity against TMPRSS2-expressing cells compared to earlier variants.
Keywords: Omicron; SARS-CoV-2; TMPRSS2; heparan sulfate.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
Conformational Dynamics and Binding Interactions of SARS-CoV-2 Spike Protein Variants: Omicron, XBB.1.9.2, and EG.5.J Chem Inf Model. 2025 Jul 28;65(14):7651-7667. doi: 10.1021/acs.jcim.5c00308. Epub 2025 Jul 11. J Chem Inf Model. 2025. PMID: 40643983 Free PMC article.
-
Safety and immunogenicity of a modified mRNA-lipid nanoparticle vaccine candidate against COVID-19: Results from a phase 1, dose-escalation study.Hum Vaccin Immunother. 2024 Dec 31;20(1):2408863. doi: 10.1080/21645515.2024.2408863. Epub 2024 Oct 18. Hum Vaccin Immunother. 2024. PMID: 39422261 Free PMC article. Clinical Trial.
-
Physical interventions to interrupt or reduce the spread of respiratory viruses.Cochrane Database Syst Rev. 2023 Jan 30;1(1):CD006207. doi: 10.1002/14651858.CD006207.pub6. Cochrane Database Syst Rev. 2023. PMID: 36715243 Free PMC article.
-
SARS-CoV-2-neutralising monoclonal antibodies to prevent COVID-19.Cochrane Database Syst Rev. 2022 Jun 17;6(6):CD014945. doi: 10.1002/14651858.CD014945.pub2. Cochrane Database Syst Rev. 2022. PMID: 35713300 Free PMC article.
References
-
- Daly JL, Simonetti B, Klein K, Chen KE, Williamson MK, Antón-Plágaro C, Shoemark DK, Simón-Gracia L, Bauer M, Hollandi R, Greber UF, Horvath P, Sessions RB, Helenius A, Hiscox JA, Teesalu T, Matthews DA, Davidson AD, Collins BM, Cullen PJ, Yamauchi Y. 2020. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 370:861–865. doi: 10.1126/science.abd3072 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
