

THAP11-mediated K48- and K63-linked ubiquitination is essential for the degradation of porcine reproductive and respiratory syndrome virus nonstructural protein 1β
- PMID: 40548980
- PMCID: PMC12185814
- DOI: 10.1007/s00018-025-05760-3
THAP11-mediated K48- and K63-linked ubiquitination is essential for the degradation of porcine reproductive and respiratory syndrome virus nonstructural protein 1β
Abstract
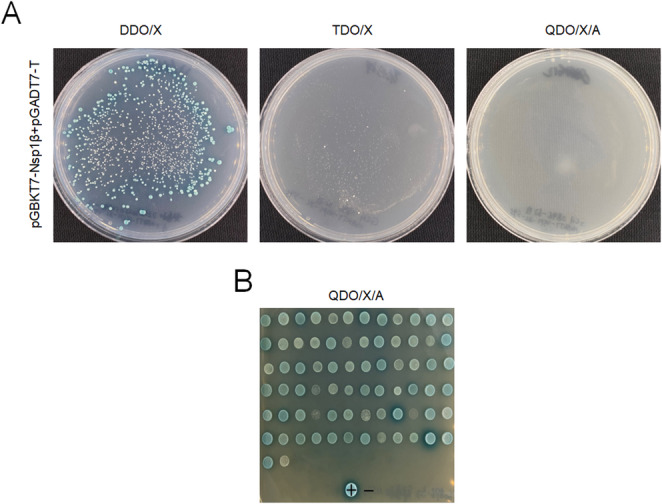
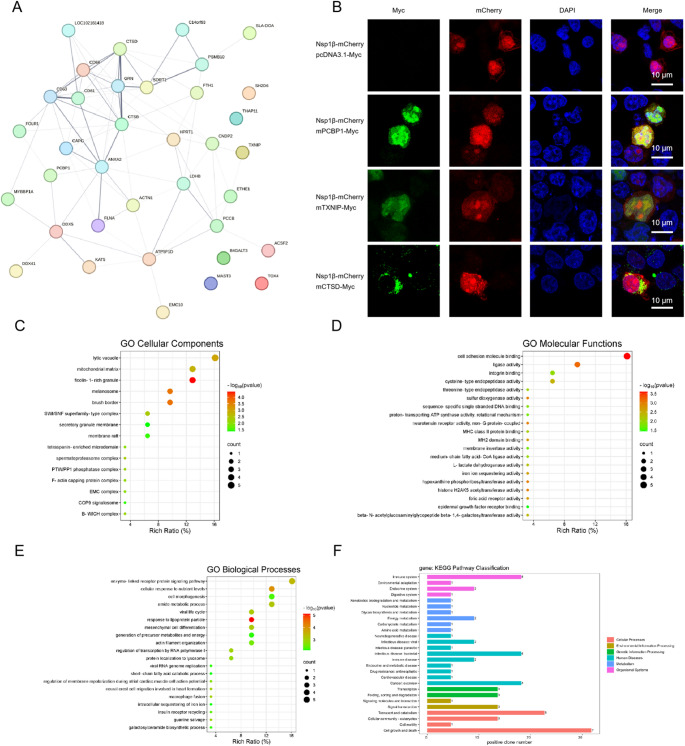
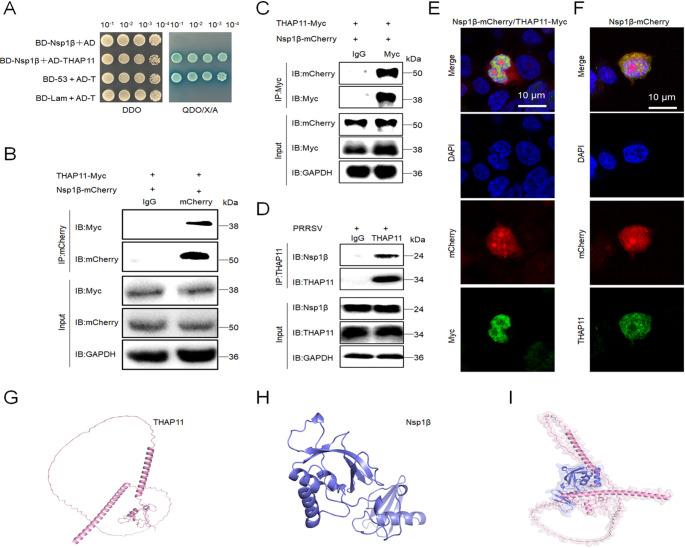
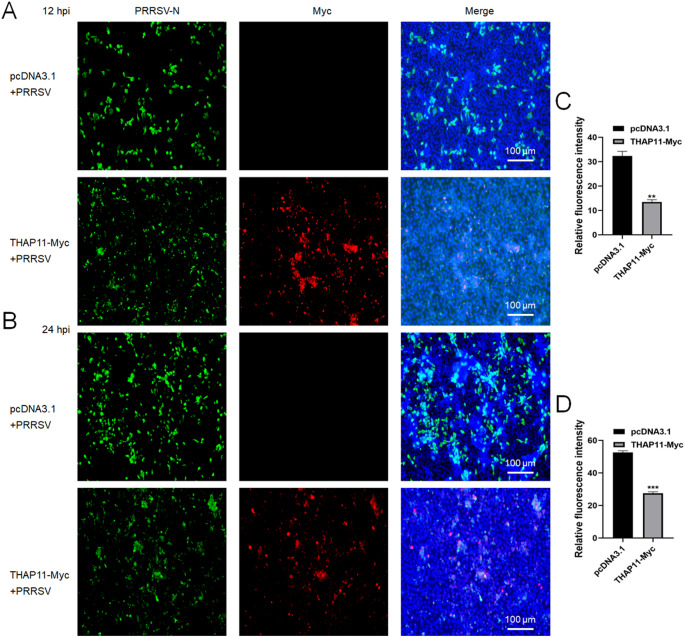
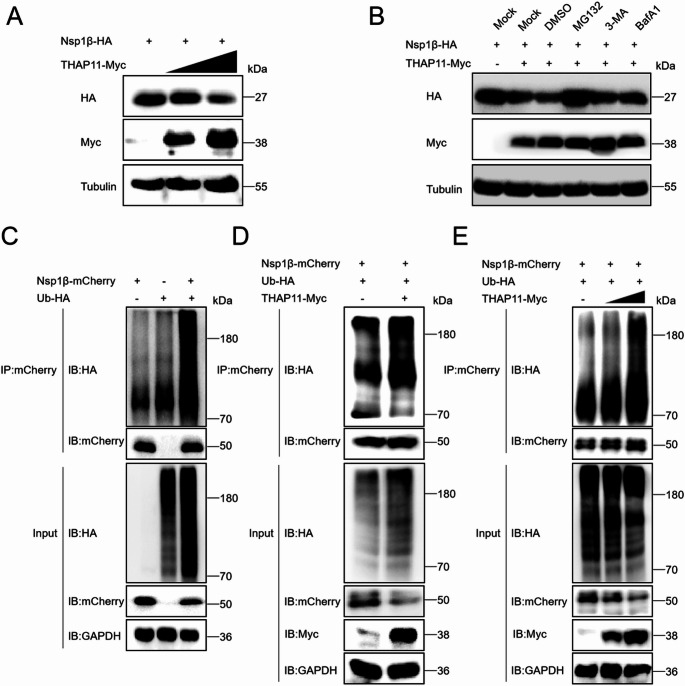
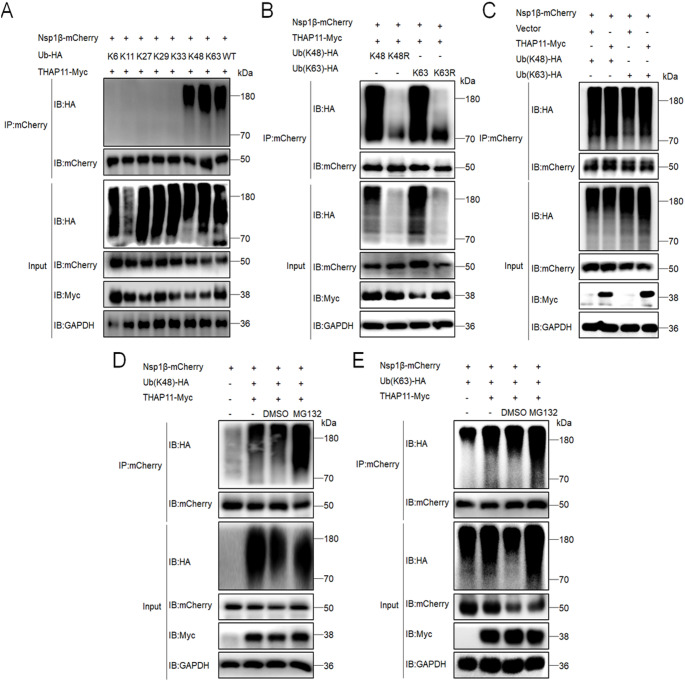
Porcine reproductive and respiratory syndrome virus (PRRSV) is a highly infectious pathogen in the global pig industry that causes significant economic losses. Owing to its rapid mutation, effective antiviral treatments or vaccines are still lacking. Therefore, it is essential to identify potential host factors that interact with PRRSV-encoded proteins. In this study, a porcine alveolar macrophage cDNA library was used to identify host proteins that interact with PRRSV nonstructural protein 1β (Nsp1β) via a yeast two-hybrid system. A total of 34 potential host factors were identified, with Thanatos-associated protein 11 (THAP11) strongly interacting with Nsp1β. These interactions were further analyzed via Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses. Co-localization of Nsp1β with THAP11, poly(rC)-binding protein 1 (PCBP1), thioredoxin-interacting protein (TXNIP), and cathepsin D (CTSD) was observed, and co-IP assays confirmed the Nsp1β-THAP11 interaction. The overexpression of THAP11 reduced PRRSV N protein accumulation, indicating an antiviral effect, whereas the silencing of THAP11 increased PRRSV replication. Furthermore, THAP11 promoted the degradation of Nsp1β by increasing K48- and K63-linked ubiquitination, thereby restricting PRRSV replication. These findings suggest that THAP11 exerts an antiviral effect by interacting with and degrading Nsp1β via the ubiquitin-proteasome system, providing insights for future PRRSV defence strategies.
Keywords: Nsp1β; PRRSV; THAP11; Virus-host interactions; Yeast two-hybrid screening.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interest: The authors declare that they have no conflicts of interest.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
