

SARS-CoV-2 nucleocapsid protein directly prevents cGAS-DNA recognition through competitive binding
- PMID: 40549905
- PMCID: PMC12232725
- DOI: 10.1073/pnas.2426204122
SARS-CoV-2 nucleocapsid protein directly prevents cGAS-DNA recognition through competitive binding
Abstract
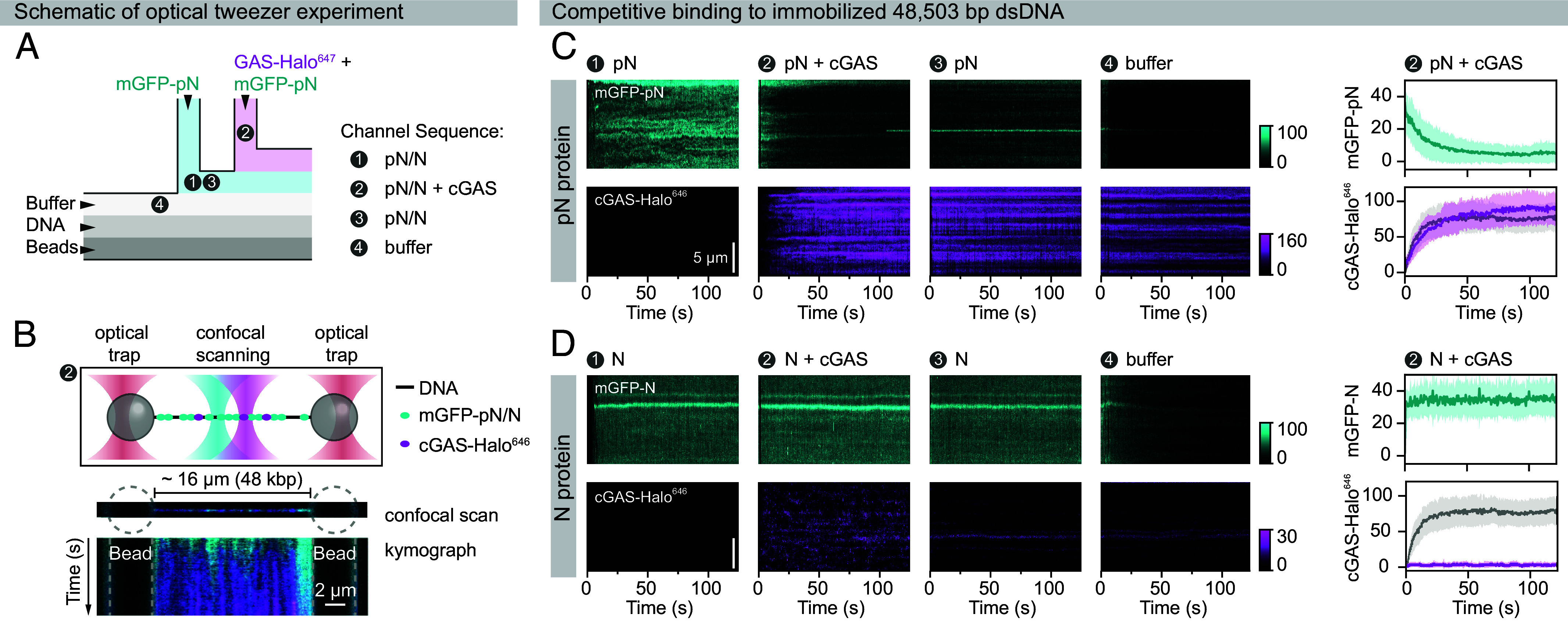
A hallmark of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection is the delayed interferon response. Interferons are typically produced upon host recognition of pathogen- or damage-associated molecular patterns, such as nucleic acids. While the mechanisms by which SARS-CoV-2 evades host recognition of its RNA are well studied, how it evades immune responses to cytosolic DNA-leaked from mitochondria or nuclei during infection-remains poorly understood. Here, we demonstrate that the SARS-CoV-2 nucleocapsid protein directly suppresses DNA sensing by cyclic guanosine monophosphate-adenosine monophosphate synthase (cGAS). Although primarily known for packaging the viral RNA genome, we uncover that the SARS-CoV-2 nucleocapsid protein also binds DNA with high affinity and competitively blocks cGAS activation. Using cell-free biochemical and biophysical approaches, including single-molecule optical tweezers, we show that the nucleocapsid protein binds to DNA at nanomolar concentrations and cocondenses with DNA at micromolar concentrations, thereby impeding stable cGAS-DNA interactions required for signal propagation. Hyperphosphorylation of the nucleocapsid protein diminishes its competitive binding capacity. Our findings reveal an unexpected role of the SARS-CoV-2 nucleocapsid protein in directly suppressing the cGAS-STING pathway, strongly suggesting that this contributes to the delayed interferon response during infection. This study raises the possibility that nucleocapsid proteins of other RNA viruses may also exhibit moonlighting functions by antagonizing host nucleic acid-sensing pathways.
Keywords: biomolecular condensates; cGAS; coronavirus; innate immune system; nucleocapsid protein.
Conflict of interest statement
Competing interests statement:A.A.H. is a founder of and consultant for Dewpoint Therapeutics Inc. The company was not involved in the work of this manuscript.
Figures
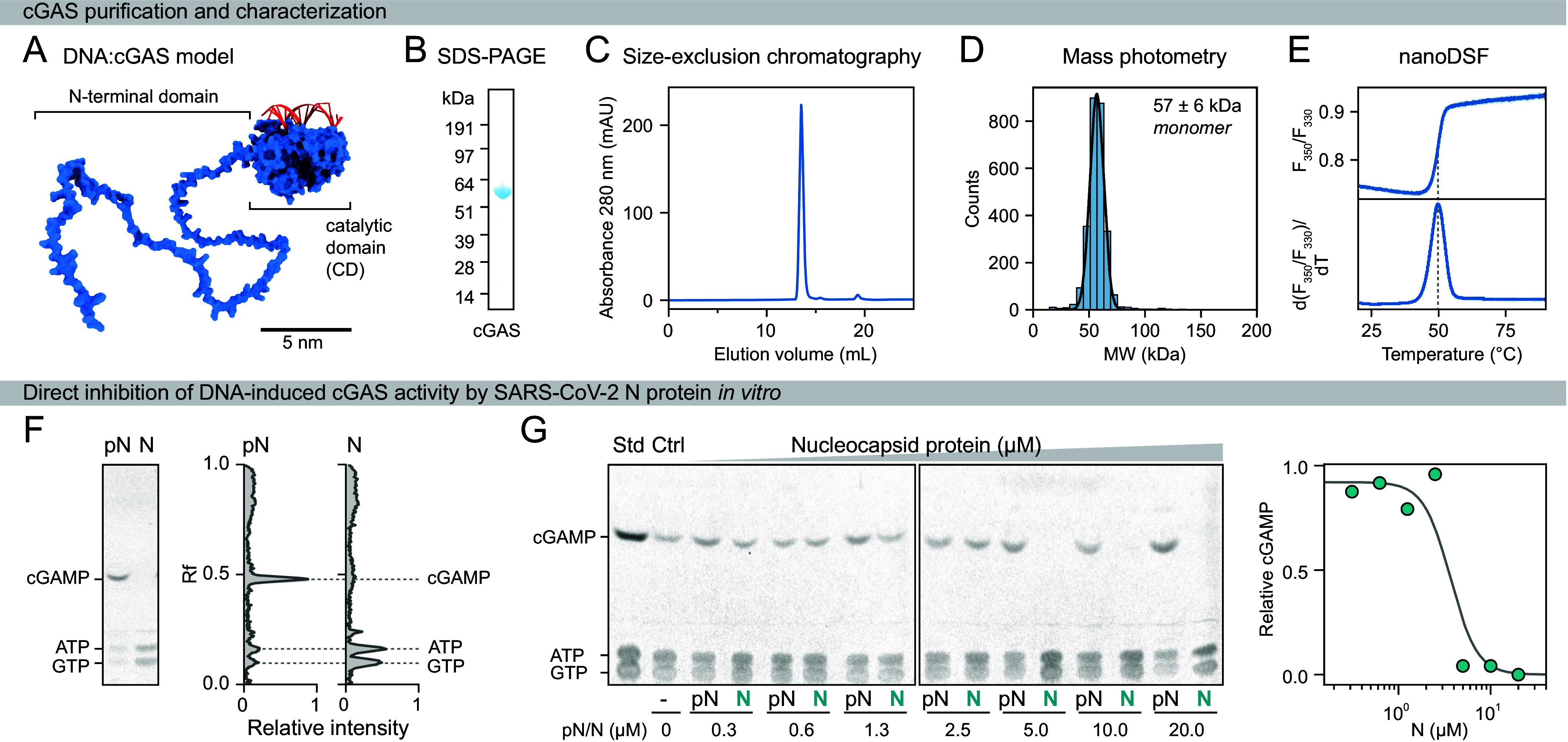
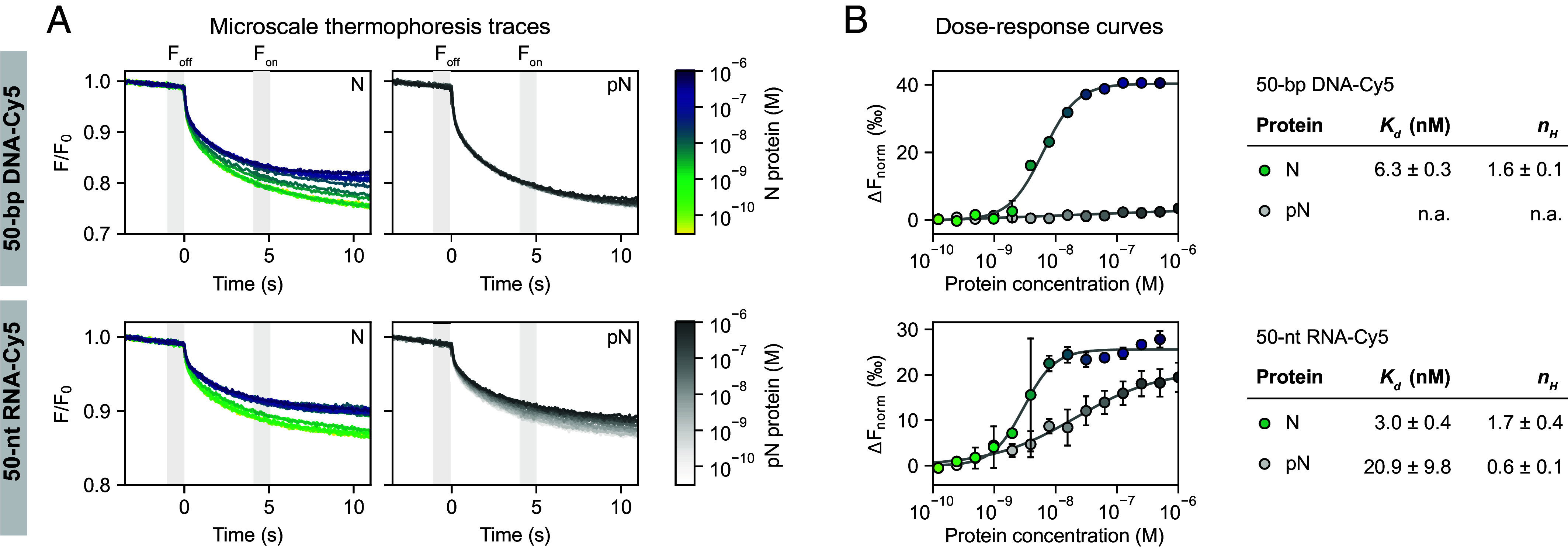
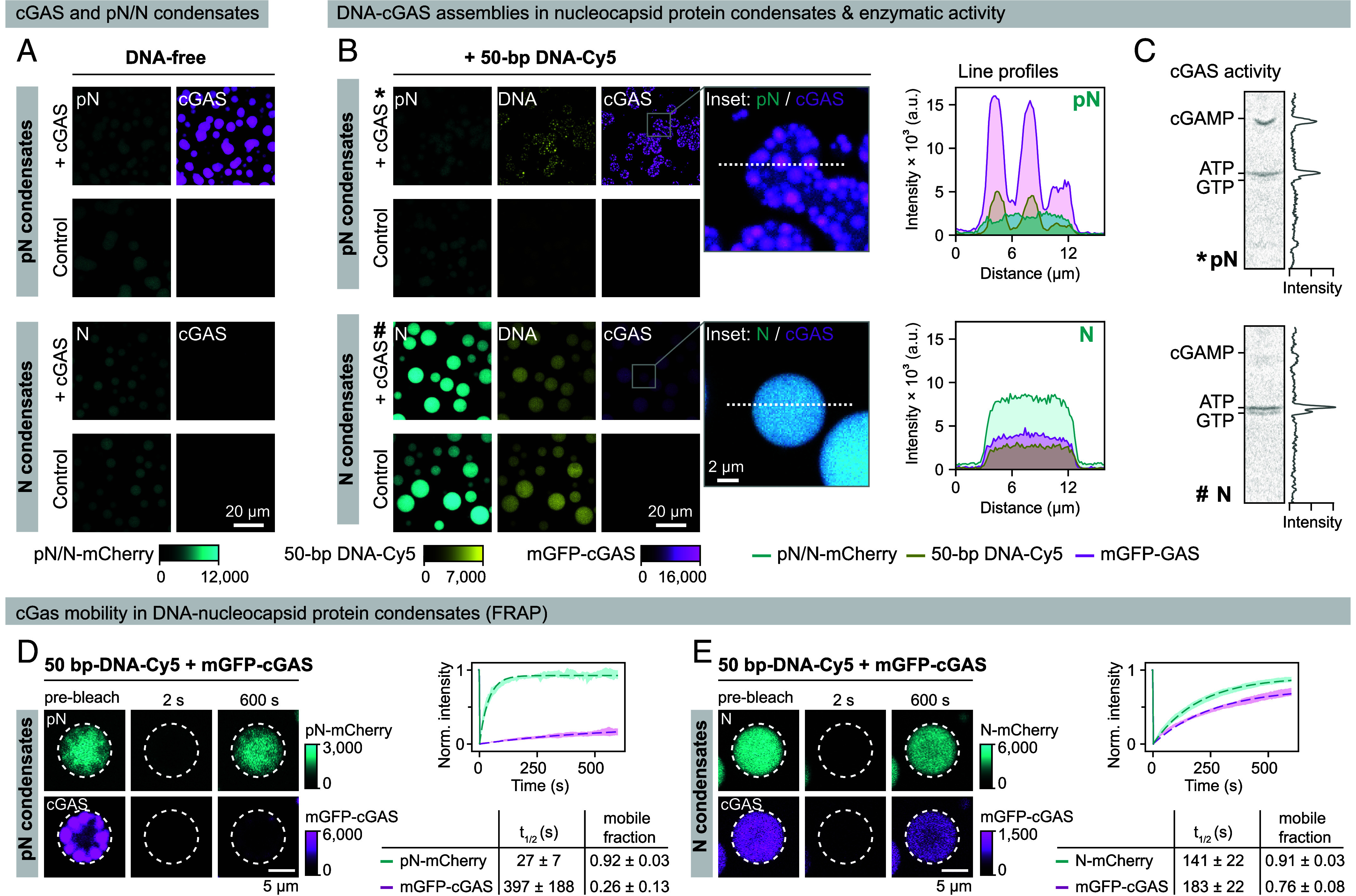
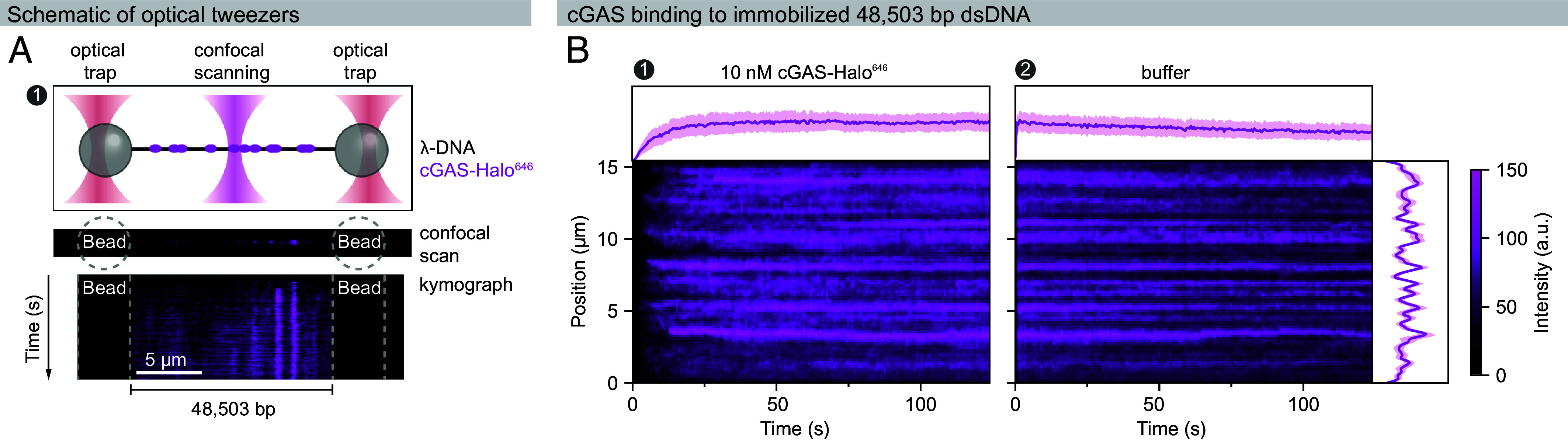
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
