

Phenotypic analysis of various Clostridioides difficile ribotypes reveals consistency among core processes
- PMID: 40552806
- PMCID: PMC12285255
- DOI: 10.1128/aem.00964-25
Phenotypic analysis of various Clostridioides difficile ribotypes reveals consistency among core processes
Abstract
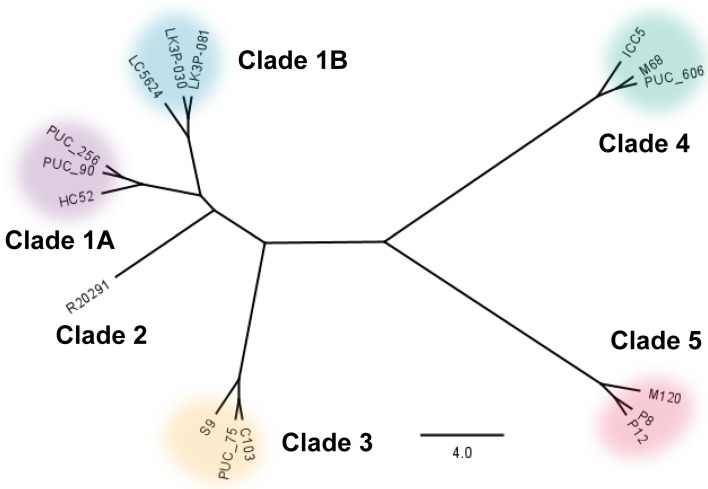
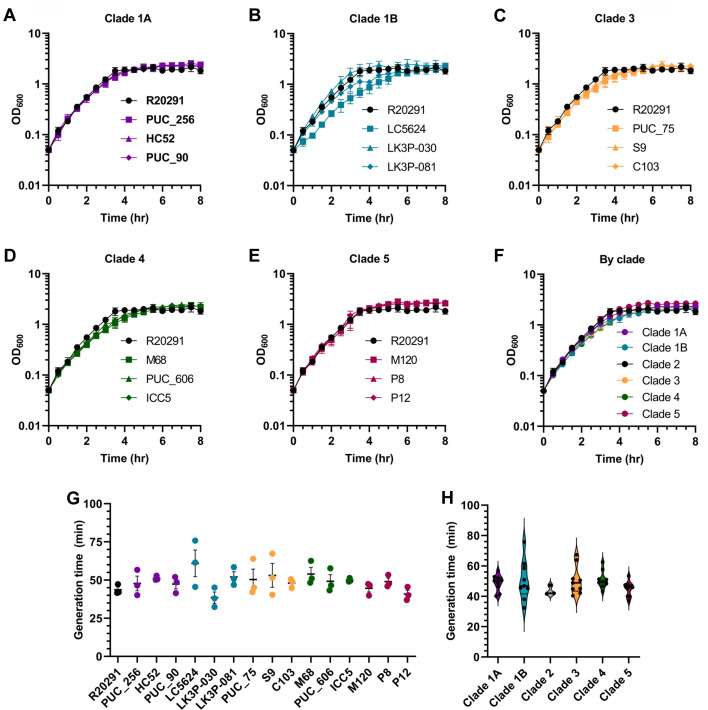
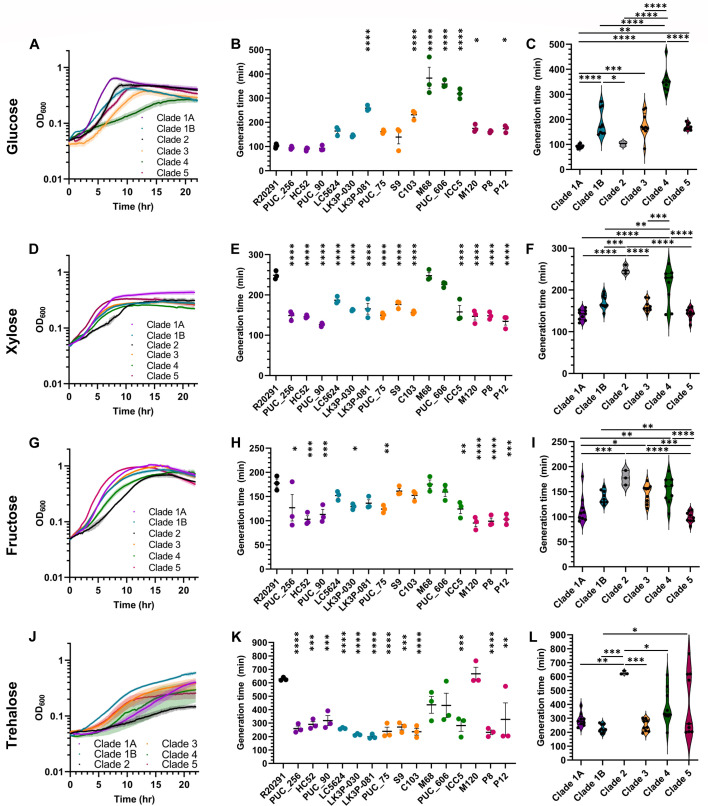
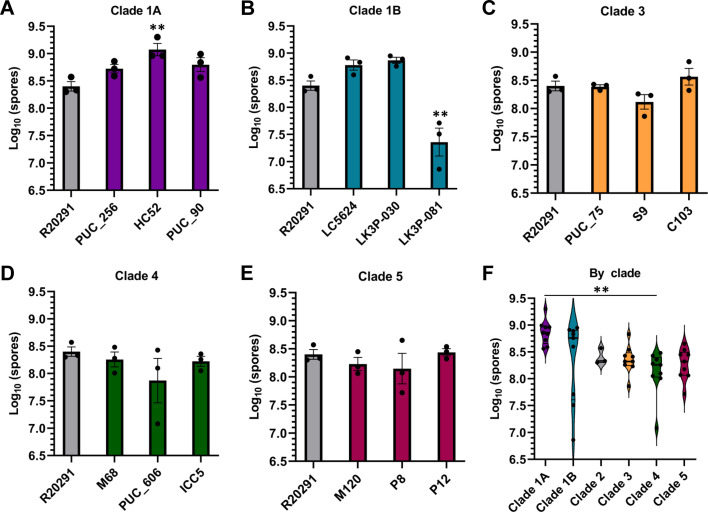
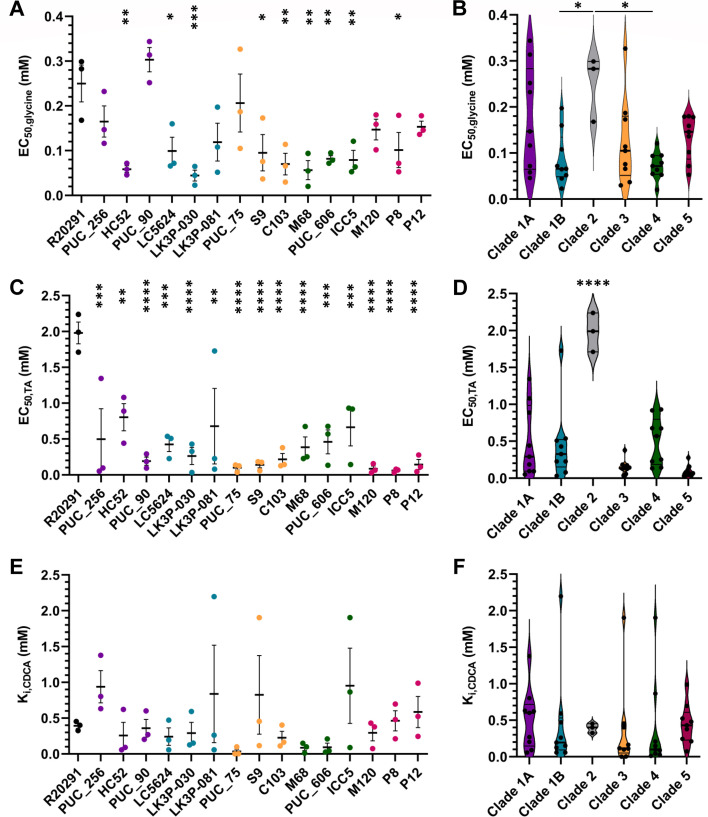
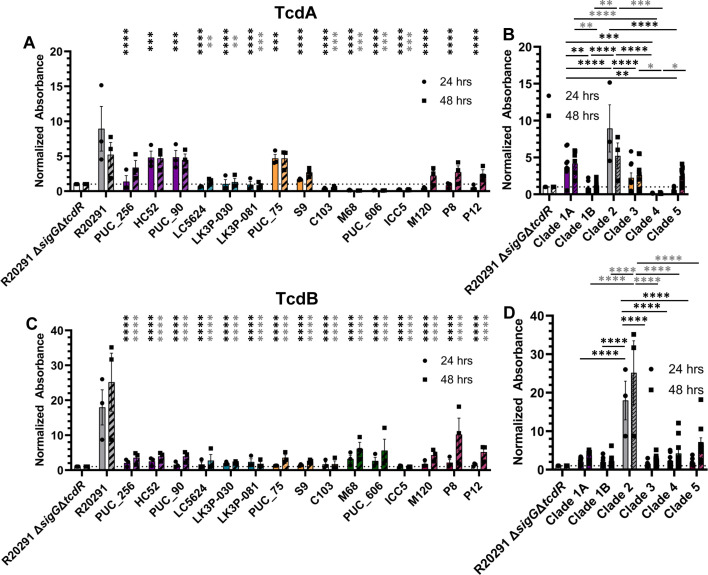
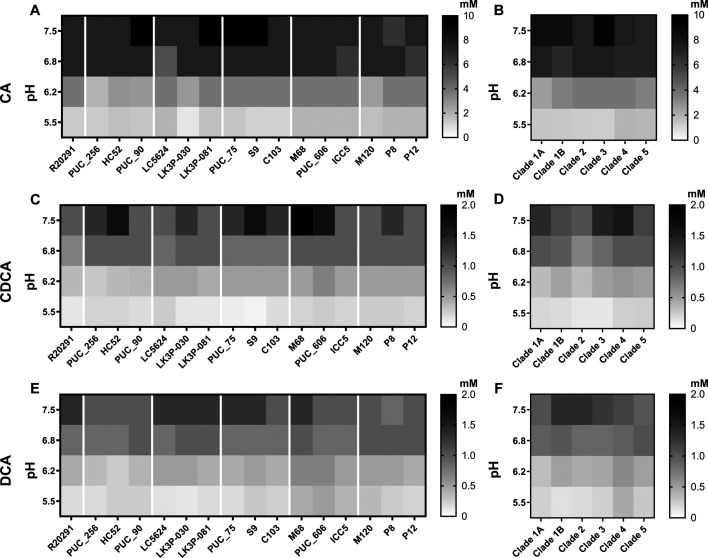
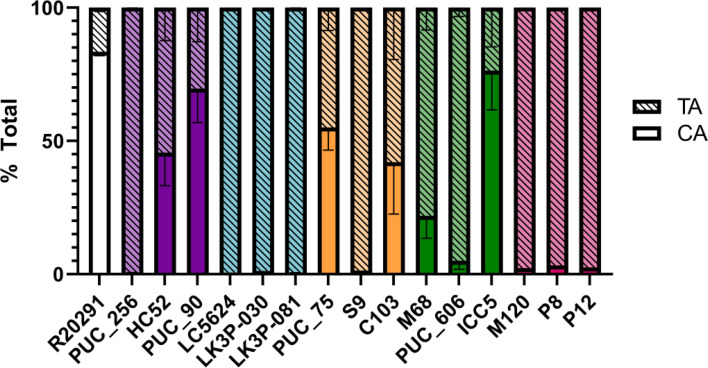
Clostridioides difficile infections (CDI) cause almost 300,000 hospitalizations per year, of which ~15%-30% are the result of recurring infections. The prevalence and persistence of CDI in hospital settings have resulted in an extensive collection of C. difficile clinical isolates and their classification, typically by ribotype. While much of the current literature focuses on one or two prominent epidemic ribotypes (e.g., RT027), recent years have seen several other ribotypes dominate the clinical landscape (e.g., RT106 and RT078). Some ribotypes are associated with severe disease and/or increased recurrence rates, but why certain ribotypes are more prominent or harmful than others remains unknown. Because C. difficile has a large, open pan-genome, this observed relationship between ribotype and clinical outcome could be a result of the genetic diversity of C. difficile. Thus, we hypothesize that the core biological processes of C. difficile are conserved across ribotypes/clades. We tested this hypothesis by observing the growth kinetics, sporulation, germination, production of toxin A and toxin B, bile acid sensitivity, bile salt hydrolase activity, and surface motility of 15 strains belonging to various ribotypes spanning each known C. difficile clade. In viewing these phenotypes across each strain, we see that core phenotypes (growth, germination, sporulation, and resistance to bile salt toxicity) are remarkably consistent across clades/ribotypes. This suggests that variations observed in the clinical setting may be due to unidentified factors in the accessory genome or due to unknown host factors.IMPORTANCEClostridioides difficile infections impact thousands of individuals every year, many of whom experience recurring infections. Clinical studies have reported an unexplained correlation between some clades/ribotypes of C. difficile and disease severity/recurrence. Here, we demonstrate that C. difficile strains across major clades/ribotypes are consistent in their core phenotypes. This suggests that such phenotypes are not responsible for variations in disease severity/recurrence and are ideal targets for the development of therapeutics meant to treat C. difficile-related infections.
Keywords: Clostridium difficile; clade; phenotypes; physiology; ribotype.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Update of
-
Phenotypic analysis of various Clostridioides difficile ribotypes reveals consistency among core processes.bioRxiv [Preprint]. 2025 Jan 10:2025.01.10.632434. doi: 10.1101/2025.01.10.632434. bioRxiv. 2025. Update in: Appl Environ Microbiol. 2025 Jul 23;91(7):e0096425. doi: 10.1128/aem.00964-25. PMID: 39829883 Free PMC article. Updated. Preprint.
References
-
- Kochan TJ, Shoshiev MS, Hastie JL, Somers MJ, Plotnick YM, Gutierrez-Munoz DF, Foss ED, Schubert AM, Smith AD, Zimmerman SK, Carlson PE Jr, Hanna PC. 2018. Germinant synergy facilitates Clostridium difficile spore germination under physiological conditions. mSphere 3:e00335-18. doi: 10.1128/mSphere.00335-18 - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
