

miR-29a-3p and TGF-β Axis in Fanconi anemia: mechanisms driving metabolic dysfunction and genome stability
- PMID: 40555815
- PMCID: PMC12187631
- DOI: 10.1007/s00018-025-05775-w
miR-29a-3p and TGF-β Axis in Fanconi anemia: mechanisms driving metabolic dysfunction and genome stability
Abstract
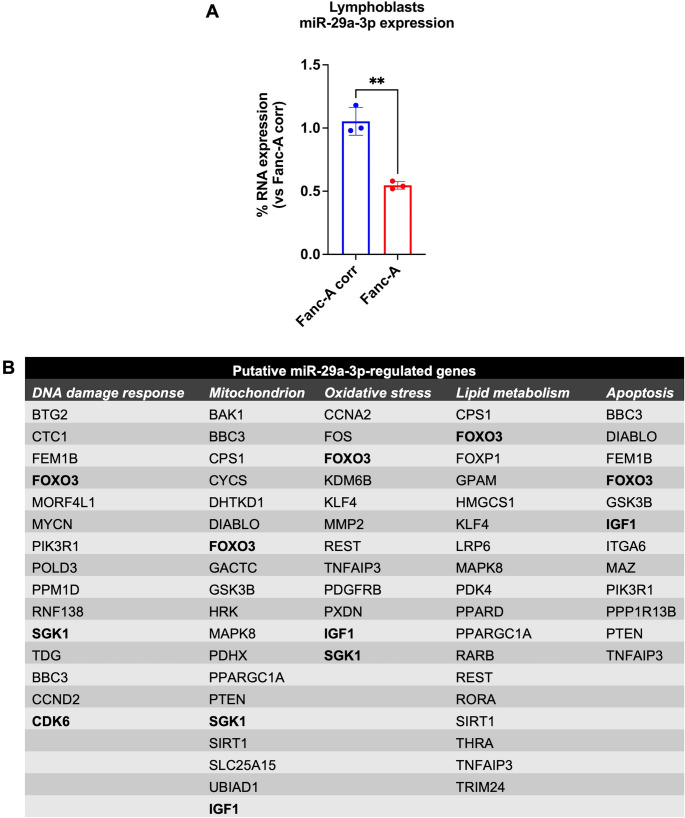
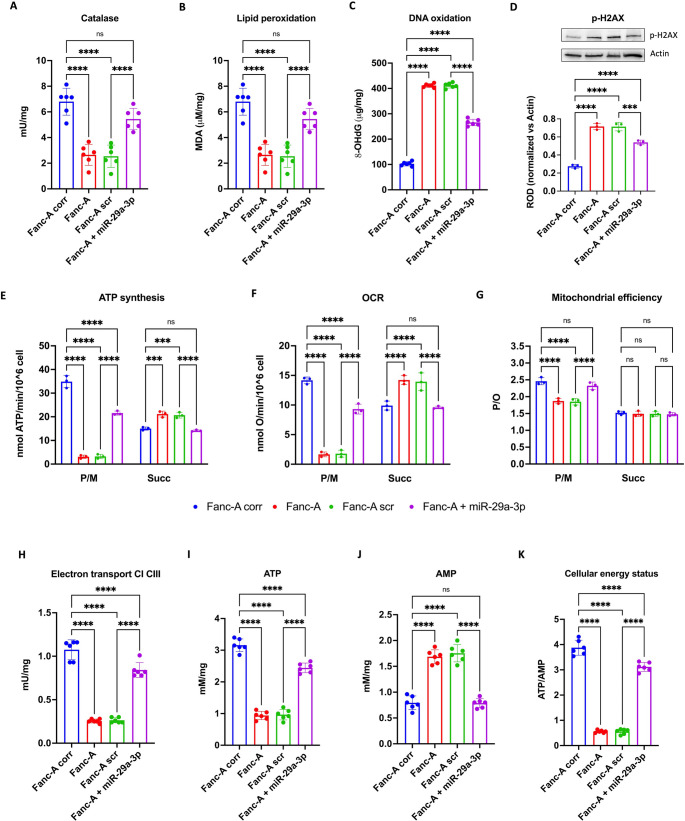
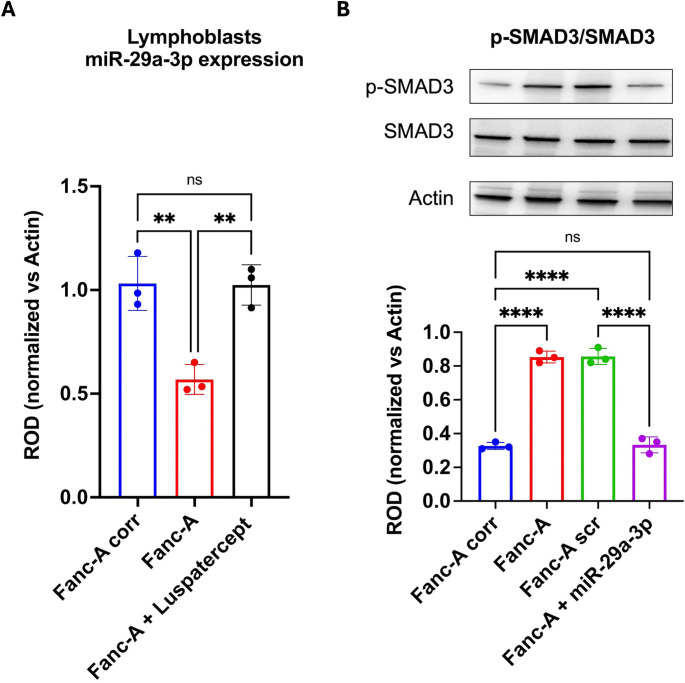
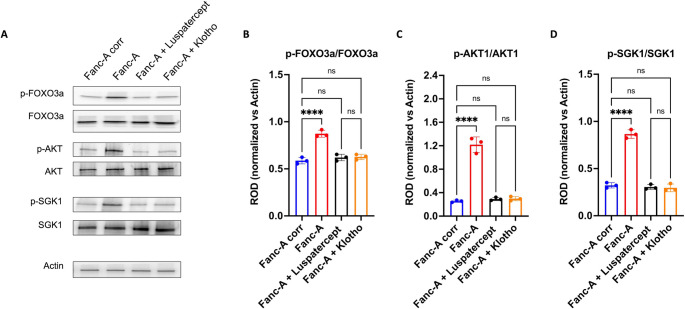
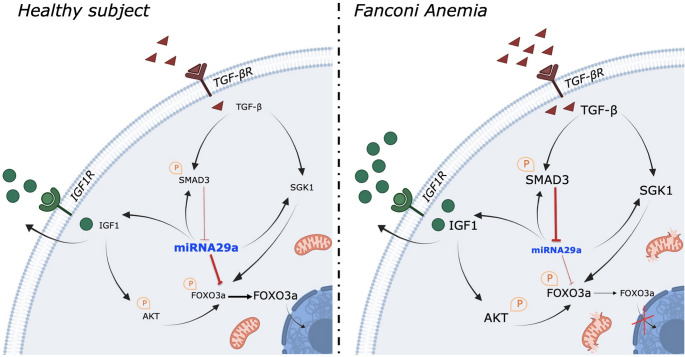
Fanconi anemia (FA) is a genetic disorder characterized by bone marrow failure and cancer predisposition. The FA cellular phenotype is marked by a defective DNA double-strand break repair. Alongside this defect, FA cells exhibit mitochondrial dysfunction and redox unbalance. In addition, FA cells display an altered microRNA profile, including miR-29a-3p, which plays a crucial role in hematopoiesis by supporting the self-renewal, lineage commitment, and differentiation of hematopoietic stem cells (HSCs). In this study, we demonstrate that miR-29a-3p is downregulated in lymphoblasts and fibroblasts mutated for the FANC-A gene, leading to hyperactivation of PI3K/AKT pathway due to the overexpression of its target genes, FOXO3, SGK1, and IGF1, and resulting in altered mitochondrial metabolism and insufficient antioxidant response. In addition, miR-29a-3p downregulation appears associated with hyperactivation of the TGF-β signal. By contrast, FA cells transfected with miR-29a-3p show an improvement in mitochondrial metabolism, oxidative stress response, and DNA damage accumulation, by inhibiting the PI3K/AKT pathway and modulating the TGF-β pathway through a feedback mechanism. In conclusion, our results highlight the central role of miR-29a-3p in FA cells, suggesting that it is a promising molecular target to address several mechanisms based on FA pathogenesis.
Keywords: DNA repair; Fanconi anemia; Inflammation; MiR-29a-3p; Mitochondrial metabolism.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors have no relevant financial or non-financial interests to disclose.
Figures



References
-
- de Winter JP, Joenje H (2009) The genetic and molecularbasis of Fanconi anemia. Mutat Res 668:11–19. 10.1016/j.mrfmmm.2008.11.004 - PubMed
-
- Grompe M, D’Andrea A (2001) Fanconi anemia and DNA repair. Hum Mol Genet 10:2253–2259. 10.1093/hmg/10.20.2253 - PubMed
-
- Yang Y-G, Herceg Z, Nakanishi K et al (2005) The Fanconi anemia group A proteinmodulateshomologousrepair of DNA double-strand breaks in mammaliancells. Carcinogenesis 26:1731–1740. 10.1093/carcin/bgi134 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
