

Remote Force Modulation of the T-Cell Receptor Reveals an NFAT-Threshold for CD4+ T-Cell Activation
- PMID: 40556346
- PMCID: PMC12188112
- DOI: 10.1002/eji.202451716
Remote Force Modulation of the T-Cell Receptor Reveals an NFAT-Threshold for CD4+ T-Cell Activation
Abstract
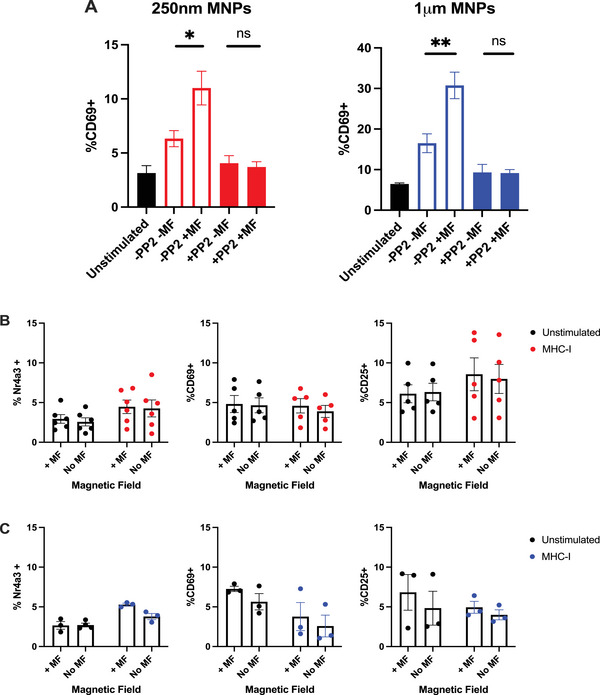
Mechano-modulation of cell surface proteins to influence cell activation has been shown as a promising new advanced therapy for regenerative medicine applications. These strategies rely on the manipulation of mechanosensitive cell surface receptors to initiate intracellular signal transduction. The cell surface receptor of T lymphocytes (TCR), which recognises peptide-MHC molecules central to driving the adaptive immune response, has recently been suggested to be mechano-responsive. Despite this advance, little is known as to whether the TCR can be mechanically modulated to achieve TCR signalling and subsequent T-cell activation, and whether these characteristics can be exploited for immunotherapies. Here, we describe a magnetic particle-based platform for mechanical modulation of the TCR and outline how this platform can be utilised to achieve CD4+ T-cell activation. We demonstrate that mechanical manipulation of the TCR induces cell surface clustering of the TCR and downstream TCR signalling, leading to eventual TCR downregulation and T-cell activation. We investigate the temporal relationship between mechanical modulation of the TCR and subsequent T-cell activation, thereby identifying that accumulation of signalling events within the NFAT pathway is required to reach the threshold required for CD4+ T-cell activation, outlining an axis which controls the CD4+ T-cell response to external mechanical cues. These findings identify how CD4+ T cells can modulate their function in response to such cues while also outlining a remote-magnetic particle-based platform that may be used for the control of T-cell responses.
© 2025 The Author(s). European Journal of Immunology published by Wiley‐VCH GmbH.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures






Similar articles
-
How to Implement Digital Clinical Consultations in UK Maternity Care: the ARM@DA Realist Review.Health Soc Care Deliv Res. 2025 May;13(22):1-77. doi: 10.3310/WQFV7425. Health Soc Care Deliv Res. 2025. PMID: 40417997 Review.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
CD6 regulates CD4 T follicular helper cell differentiation and humoral immunity during murine coronavirus infection.J Virol. 2025 Jan 31;99(1):e0186424. doi: 10.1128/jvi.01864-24. Epub 2024 Dec 16. J Virol. 2025. PMID: 39679790 Free PMC article.
-
Assessing the comparative effects of interventions in COPD: a tutorial on network meta-analysis for clinicians.Respir Res. 2024 Dec 21;25(1):438. doi: 10.1186/s12931-024-03056-x. Respir Res. 2024. PMID: 39709425 Free PMC article. Review.
-
Eliciting adverse effects data from participants in clinical trials.Cochrane Database Syst Rev. 2018 Jan 16;1(1):MR000039. doi: 10.1002/14651858.MR000039.pub2. Cochrane Database Syst Rev. 2018. PMID: 29372930 Free PMC article.
Cited by
-
Mechanical interactions impact the functions of immune cells and their application in immunoengineering.Adv Ther (Weinh). 2025 Jun 25:e00067. doi: 10.1002/adtp.202500067. Online ahead of print. Adv Ther (Weinh). 2025. PMID: 40881797
References
-
- Krogsgaard M., Li Q.‐J., Sumen C., Huppa J. B., Huse M., and Davis M. M., “Agonist/Endogenous Peptide‐MHC Heterodimers Drive T Cell Activation and Sensitivity,” Nature 434 (2005): 238–243. - PubMed
-
- Stefanová I., Dorfman J. R., and Germain R. N., “Self‐recognition Promotes the Foreign Antigen Sensitivity of Naive T Lymphocytes,” Nature 420 (2002): 429–434. - PubMed
-
- van der Merwe P. A., “Do T Cell Receptors Do It Alone?,” Nature Immunology 3 (2002): 1122–1123. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
