

High Maternal Glycine Levels Increase the Risk of Developing Atrial Septal Defect in the Offspring
- PMID: 40561639
- PMCID: PMC12226122
- DOI: 10.1016/j.jacbts.2025.05.006
High Maternal Glycine Levels Increase the Risk of Developing Atrial Septal Defect in the Offspring
Abstract
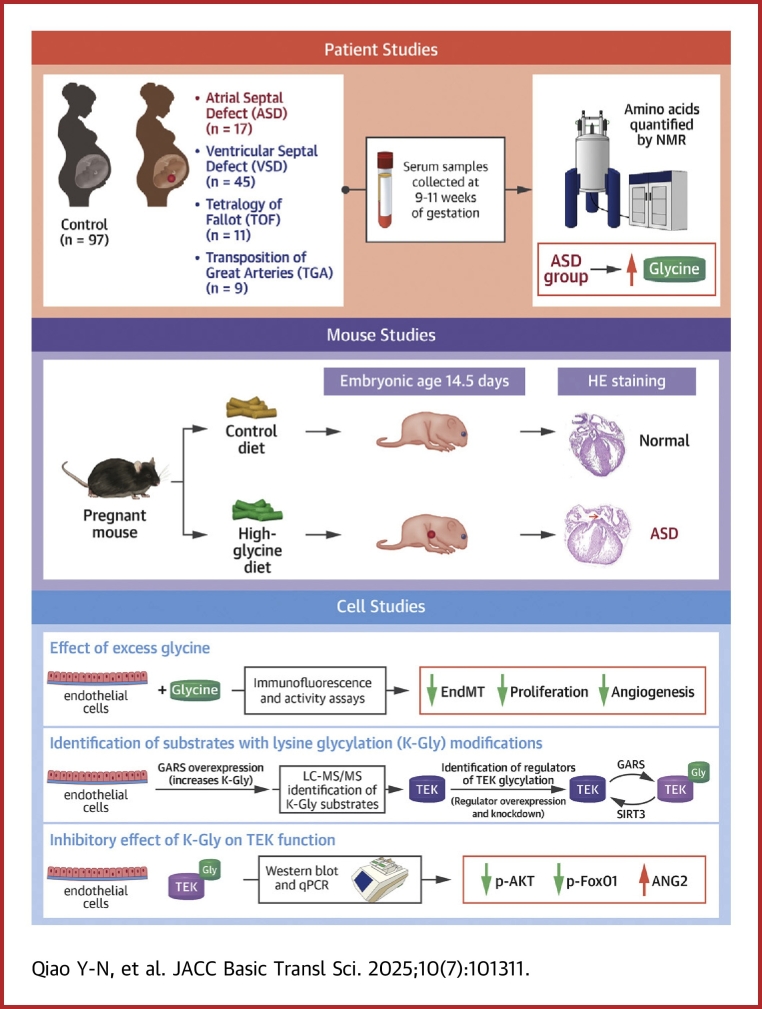
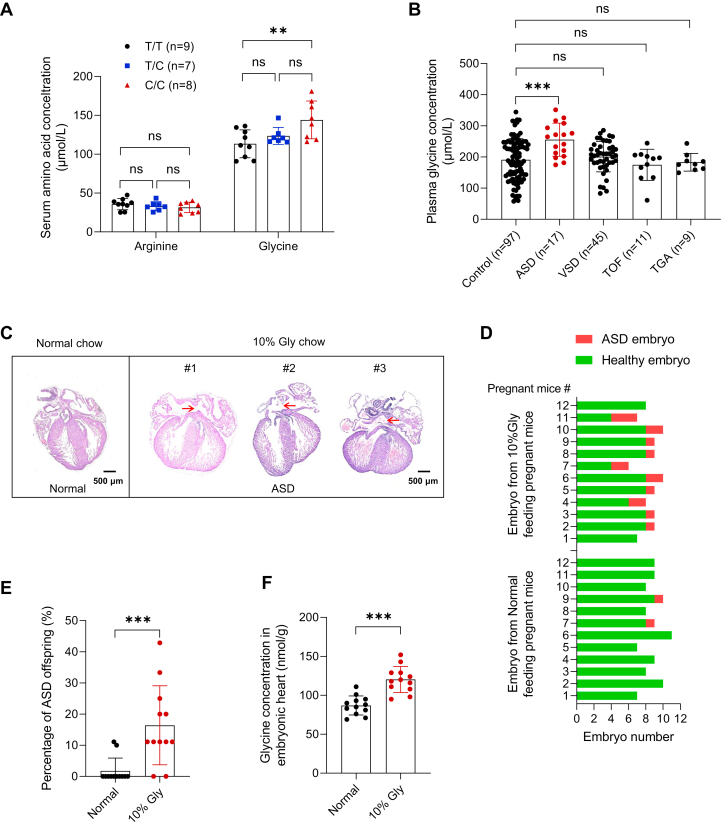
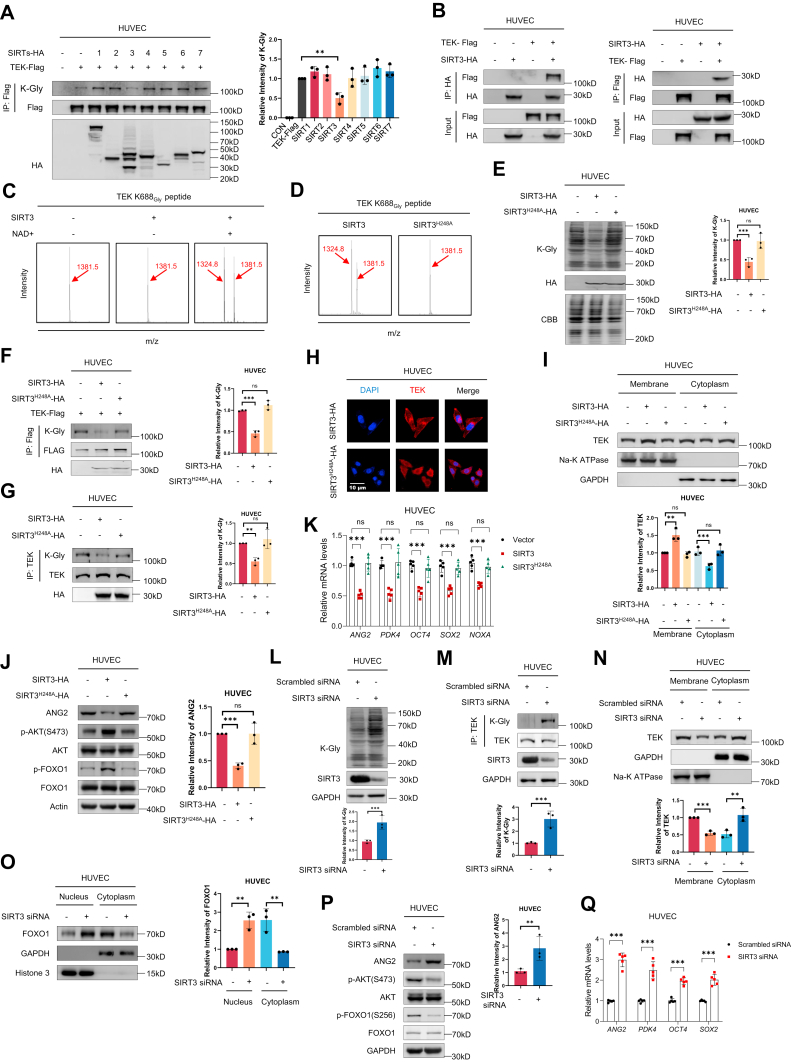
Amino acid imbalance is linked to increased congenital heart disease risk. Here, we found women carrying rs2545801 C/C genotypes exhibited increased glycine levels and increased risk for atrial septal defects (ASDs) in their offspring. Elevated maternal glycine levels during the first trimester were correlated with a higher ASD risk in the offspring. Additionally, feeding pregnant mice with high-glycine chow increased ASD risk in their offspring. Mechanistically, elevated maternal glycine led to increased lysine-glycylation of lysine-688 within the TEK receptor tyrosine kinase and inhibited TEK-PI3K-AKT/FOXO1 signaling in cardiac endothelial cells. These findings indicate that lysine-glycylation exerts teratogenic effects and may be a target for ASD intervention.
Keywords: TEK; atrial septal defects; glycine; maternal single-nucleotide variation; pregnancy serum.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Funding Support and Author Disclosures This work was supported by the grants from National Natural Science Foundation of China (82330048, 82170236, 81700212, 32000895, 32370824, 82300428), Program of Shanghai Academic Research Leader (21XD1421700), Innovation Program of the Shanghai Municipal Education Commission (2023ZKZD24), and Innovative research team of high-level local universities in Shanghai (SHSMU-ZDCX20211100). The authors have reported that they have no relationships relevant to the contents of this paper to disclose.
Figures



Similar articles
-
The Effect of Maternal Choline Intake on Offspring Cognition in Adolescence: Protocol for a 14-year Follow-Up of a Randomized Controlled Feeding Trial.JMIR Res Protoc. 2025 Jul 11;14:e73508. doi: 10.2196/73508. JMIR Res Protoc. 2025. PMID: 40644695 Free PMC article.
-
Is maternal smoking during pregnancy associated with an increased risk of congenital heart defects among offspring? A systematic review and meta-analysis of observational studies.J Matern Fetal Neonatal Med. 2017 Mar;30(6):645-657. doi: 10.1080/14767058.2016.1183640. Epub 2016 May 16. J Matern Fetal Neonatal Med. 2017. PMID: 27126055
-
Antiretrovirals for reducing the risk of mother-to-child transmission of HIV infection.Cochrane Database Syst Rev. 2011 Jul 6;(7):CD003510. doi: 10.1002/14651858.CD003510.pub3. Cochrane Database Syst Rev. 2011. PMID: 21735394
-
Influenza and congenital anomalies: a systematic review and meta-analysis.Hum Reprod. 2014 Apr;29(4):809-23. doi: 10.1093/humrep/det455. Epub 2013 Dec 22. Hum Reprod. 2014. PMID: 24365800
-
Maternal Serum Folate During Pregnancy and Congenital Heart Disease in Offspring.JAMA Netw Open. 2024 Oct 1;7(10):e2438747. doi: 10.1001/jamanetworkopen.2024.38747. JAMA Netw Open. 2024. PMID: 39388179 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
