

Integrated proteomic and targeted Next Generation Sequencing reveal relevant heterogeneity in lower-grade meningioma and ANXA3 as a new target in NF2 mutated meningiomas
- PMID: 40562609
- PMCID: PMC12278414
- DOI: 10.1016/j.ebiom.2025.105814
Integrated proteomic and targeted Next Generation Sequencing reveal relevant heterogeneity in lower-grade meningioma and ANXA3 as a new target in NF2 mutated meningiomas
Abstract
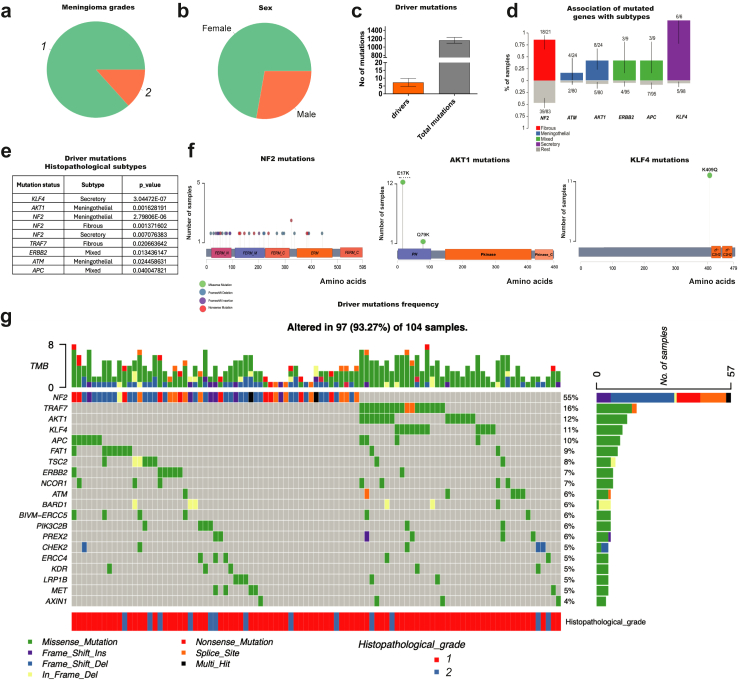
Background: Meningiomas, the most common primary brain tumours, are classified by the World Health Organization (WHO) into grades 1, 2, and 3. Some grade 1 tumours exhibit increased clinical aggressiveness, with the biallelic mutation of NF2 being the most frequently reported.
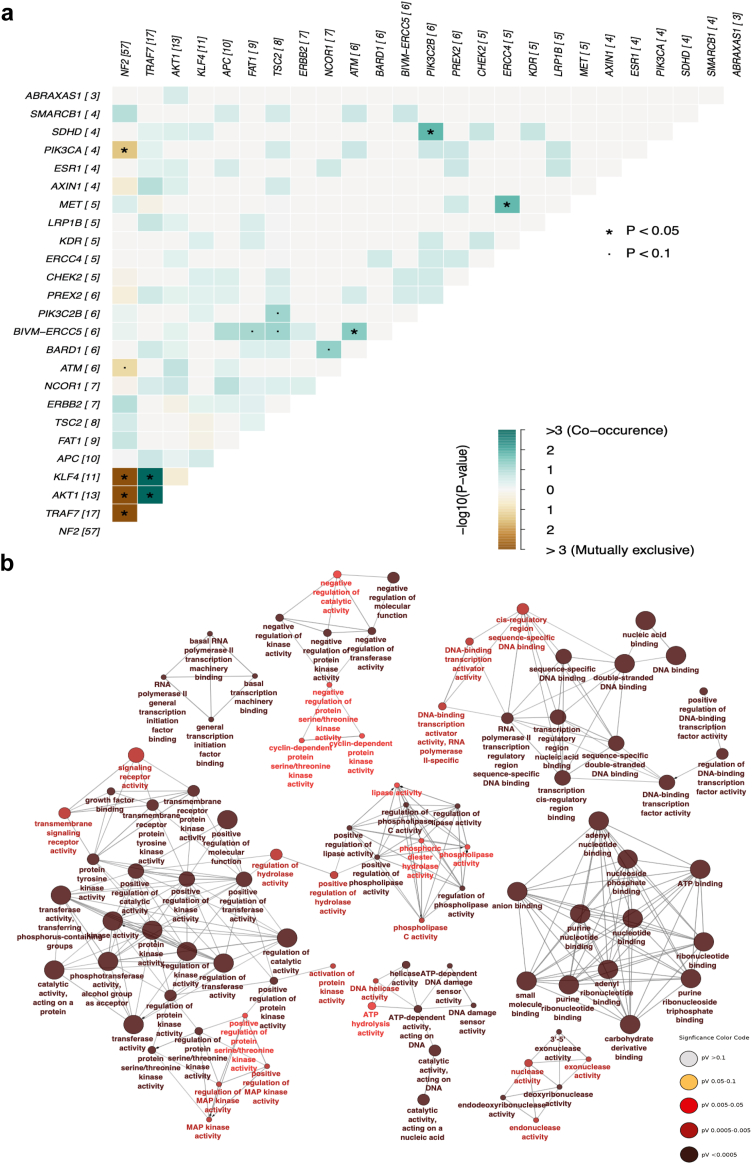
Methods: In our study, we analysed the most common driver mutations (NF2, AKT1, KLF4, and TRAF7) in meningioma by genomics describing co-occurrences and new mutations. Furthermore, tumour tissue bearing the driver mutations was analysed by proteomics. The relevance of the specific target found in the most common driver mutation in meningiomas (NF2) was validated in vitro using both lower and higher-grade meningioma and further, the higher-grade meningioma was analysed in vivo using an NOD scid gamma (NSG) mouse model.
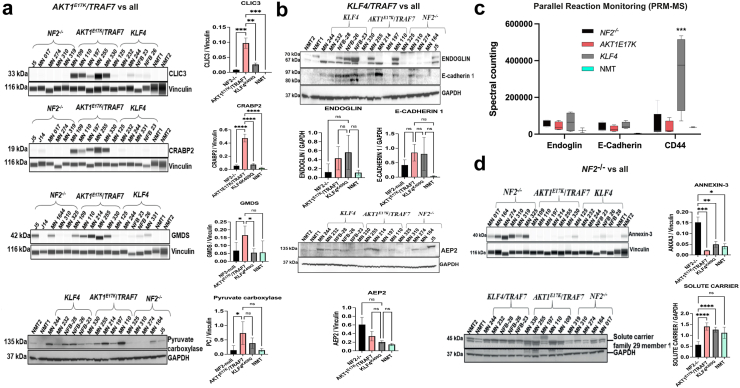
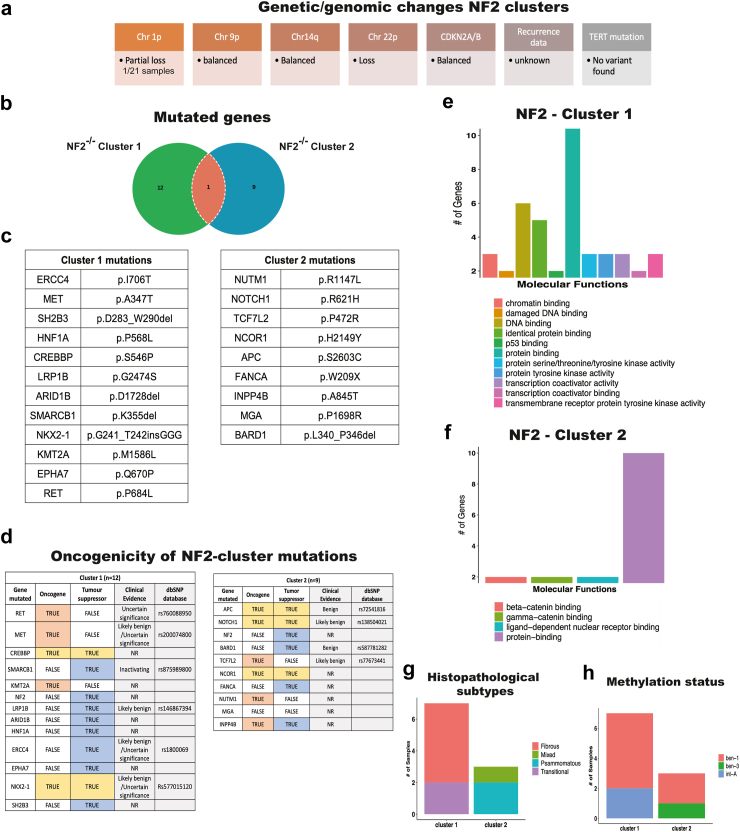
Findings: Our genomic data revealed co-occurrences of non-NF2 mutations in lower-grade meningiomas, suggesting synergistic effects supporting tumour growth. NF2-/- meningiomas showed distinct proteomic clustering, with different mutations found in these clusters. Additionally, proteomics identified Annexin-3 (ANXA3) upregulated in NF2-/- meningioma. Its role in proliferation was confirmed in grade 1 and subsequently grade 3 tumours in vitro and with abolished growth when knocked down in a meningioma mouse model.
Interpretation: These findings highlight new targets in different meningioma backgrounds, presenting ANXA3 as a potential therapeutic target for meningioma treatment.
Funding: This work was funded by the Brain Tumour Centre of Excellence.
Keywords: AKT1; ANXA3; KLF4; Meningioma; NF2; TRAF7.
Copyright © 2025 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interest. COH in 2022 received consulting fees from Recursion pharma. COH is member of the International Consortium of Meningioma. This is an academic consortium.
Figures



References
-
- Sahm F., Schrimpf D., Stichel D., et al. DNA methylation-based classification and grading system for meningioma: a multicentre, retrospective analysis. Lancet Oncol. 2017;18:682–694. - PubMed
-
- Parada C.A., Osbun J.W., Busald T., et al. Phosphoproteomic and kinomic signature of clinically aggressive grade I (1.5) meningiomas reveals Rb1 signaling as a novel mediator and biomarker. Clin Cancer Res. 2020;26:193–205. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
