

ETMR stem-like state and chemo-resistance are supported by perivascular cells at single-cell resolution
- PMID: 40562749
- PMCID: PMC12198369
- DOI: 10.1038/s41467-025-60442-9
ETMR stem-like state and chemo-resistance are supported by perivascular cells at single-cell resolution
Abstract
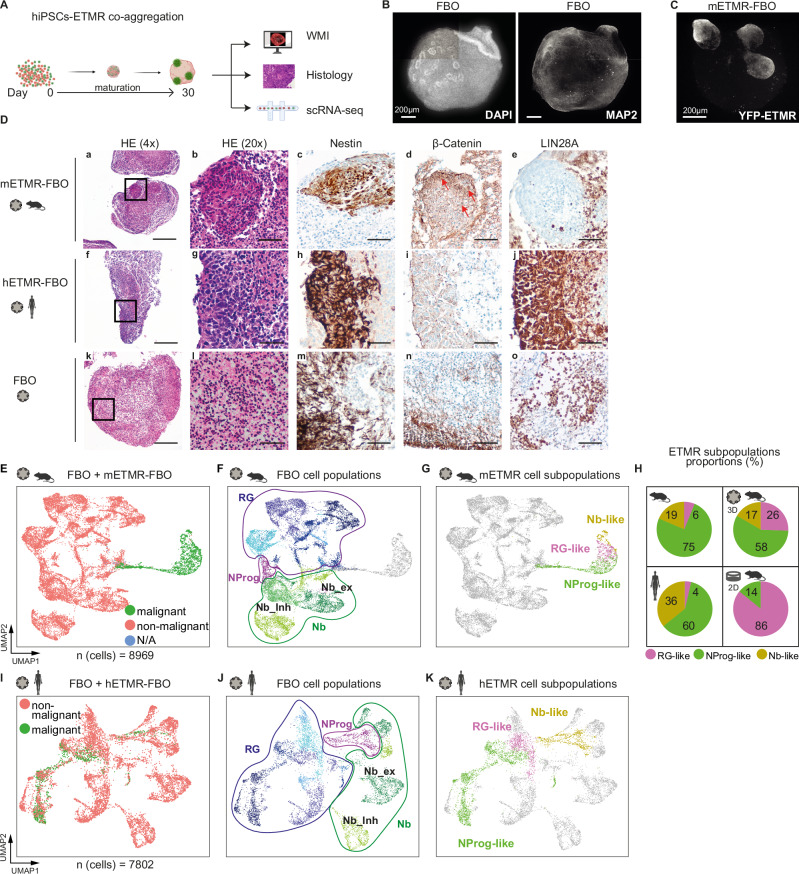
Embryonal tumor with multilayered rosettes (ETMR) is a lethal embryonal brain tumor entity. To investigate the intratumoral heterogeneity and cellular communication in the tumor microenvironment (TME), we analyze in this work single-cell RNA sequencing of about 250,000 cells of primary human and murine ETMR, in vitro cultures, and a 3D forebrain organoid model of ETMR, supporting the main findings with immunohistochemistry and spatial transcriptomics of human tumors. We characterize three distinct malignant ETMR subpopulations - RG-like, NProg-like and NB-like - positioned within a putative neurodevelopmental hierarchy. We reveal PDGFRβ+ pericytes as key communication partners in the TME, contributing to stem cell signaling through extracellular matrix-mediated interactions with tumor cells. PDGF signaling is upregulated in chemoresistant RG-like cells in vivo and plays a role in recruiting pericytes to ETMR TME by finalizing a signaling cascade which promotes the differentiation of non-malignant radial glia cells, derived from our 3D model, into pericyte-like cells. Selective PDGFR-inhibition blocked the lineage differentiation into pericytes in vitro and reduced the tumor cell population in vivo. Targeting ETMR-pericyte interactions in the TME presents a promising therapeutic approach.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing Interests.
Figures







References
-
- Korshunov, A. et al. Focal genomic amplification at 19q13.42 comprises a powerful diagnostic marker for embryonal tumors with ependymoblastic rosettes. Acta Neuropathol.120, 253–260 (2010). - PubMed
-
- Khan S. et al. Clinical phenotypes and prognostic features of embryonal tumours with multi-layered rosettes: a Rare Brain Tumor Registry study. Lancet Child Adolesc. Health5, 800–813 (2021). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
