

Transcriptomic Analysis of the Liver Redox Response During Food-Anticipatory Activity Under a Time-Restricted Feeding Protocol in Rats
- PMID: 40563286
- PMCID: PMC12189122
- DOI: 10.3390/antiox14060649
Transcriptomic Analysis of the Liver Redox Response During Food-Anticipatory Activity Under a Time-Restricted Feeding Protocol in Rats
Abstract
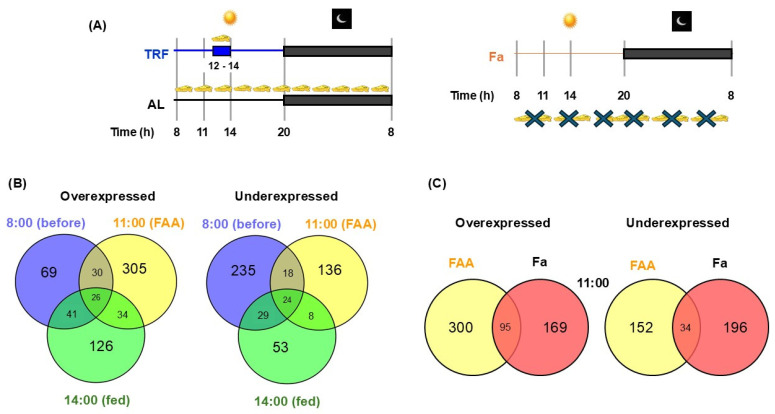
Daytime-restricted feeding (TRF) exerts outstanding effects on circadian physiology, nutrient utilization, and energy metabolism. Limiting feeding access to two hours during the daytime (12:00-14:00 h) for three weeks promotes food-anticipatory activity (FAA). FAA encompasses not only behaviors related to meal expectations but also includes diurnal fluctuations in liver metabolic responses, including distinct redox handling. Hepatic microarray profiles of genes associated with redox response processes were analyzed at three crucial time points: at the beginning of the light period or before FAA (08:00 h), during the expression of FAA (11:00 h), and after feeding (14:00 h). Data on fasting and nutrient processing were integrated, whereas circadian implications were extrapolated by comparing the TRF transcriptional output with a one-day fasting group. Transcripts of redox reactions, such as reactive oxygen species (ROS) generation, antioxidant defenses, NAD+/NADH equilibrium, and glutathione, hydrogen peroxide (H2O2), arginine, nitric oxide (NO), and hydrogen sulfide (H2S) metabolism, were analyzed. Results showed a decline in antioxidant defenses at 08:00 h, followed by a burst of pro-oxidant reactions, preparation of glutathione metabolism factors, and a tendency to decrease H2O2 and increase NO and H2S during the FAA. Most of the findings observed during the FAA were absent in response to one-day fasting. Hence, TRF involves concerted and sequential responses in liver pro-oxidant and antioxidant reactions, facilitating a redox-related circadian control that optimizes the metabolic utilization of nutrients, which differs from a response to a simple fast-feed cycle.
Keywords: food-anticipatory activity; metabolism; redox; time-restricted feeding.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures










Similar articles
-
Antibiotic-induced Dysbiosis of Gut Microbiota Increases Food Motivation and Anticipatory Activity Under a Time-restricted Feeding Protocol.J Biol Rhythms. 2025 Aug 14:7487304251359349. doi: 10.1177/07487304251359349. Online ahead of print. J Biol Rhythms. 2025. PMID: 40808497
-
Antiretrovirals for reducing the risk of mother-to-child transmission of HIV infection.Cochrane Database Syst Rev. 2011 Jul 6;(7):CD003510. doi: 10.1002/14651858.CD003510.pub3. Cochrane Database Syst Rev. 2011. PMID: 21735394
-
Nutritional labelling for healthier food or non-alcoholic drink purchasing and consumption.Cochrane Database Syst Rev. 2018 Feb 27;2(2):CD009315. doi: 10.1002/14651858.CD009315.pub2. Cochrane Database Syst Rev. 2018. PMID: 29482264 Free PMC article.
-
Antiretrovirals for reducing the risk of mother-to-child transmission of HIV infection.Cochrane Database Syst Rev. 2007 Jan 24;(1):CD003510. doi: 10.1002/14651858.CD003510.pub2. Cochrane Database Syst Rev. 2007. Update in: Cochrane Database Syst Rev. 2011 Jul 06;(7):CD003510. doi: 10.1002/14651858.CD003510.pub3. PMID: 17253490 Updated.
-
Intravenous magnesium sulphate and sotalol for prevention of atrial fibrillation after coronary artery bypass surgery: a systematic review and economic evaluation.Health Technol Assess. 2008 Jun;12(28):iii-iv, ix-95. doi: 10.3310/hta12280. Health Technol Assess. 2008. PMID: 18547499
References
-
- García-Gaytán A.C., Miranda-Anaya M., Turrubiate I., López-De Portugal L., Bocanegra-Botello G.N., López-Islas A., Díaz-Muñoz M., Méndez I. Synchronization of the circadian clock by time-restricted feeding with progressive increasing calorie intake. Resemblances and differences regarding a sustained hypocaloric restriction. Sci. Rep. 2020;10:10036. doi: 10.1038/s41598-020-66538-0. - DOI - PMC - PubMed
-
- Aguilar-Roblero R., Díaz-Muñoz M. Chronostatic adaptations in the liver to restricted feeding: The FEO as an emergent oscillator. Sleep Biol. Rhythm. 2010;8:9–17. doi: 10.1111/j.1479-8425.2009.00415.x. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
