

KRAS and NRF2 drive metabolic reprogramming in pancreatic cancer cells: the influence of oxidative and nitrosative stress
- PMID: 40567499
- PMCID: PMC12187747
- DOI: 10.3389/fcell.2025.1547582
KRAS and NRF2 drive metabolic reprogramming in pancreatic cancer cells: the influence of oxidative and nitrosative stress
Erratum in
-
Correction: KRAS and NRF2 drive metabolic reprogramming in pancreatic cancer cells: the influence of oxidative and nitrosative stress.Front Cell Dev Biol. 2025 Aug 8;13:1661525. doi: 10.3389/fcell.2025.1661525. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40861269 Free PMC article.
Abstract
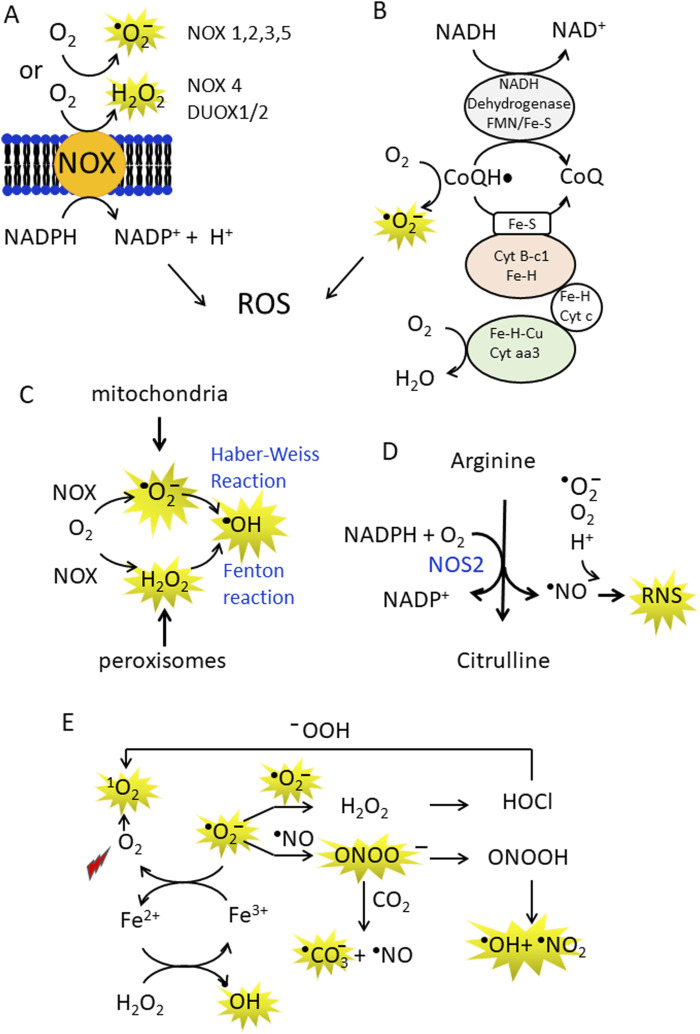
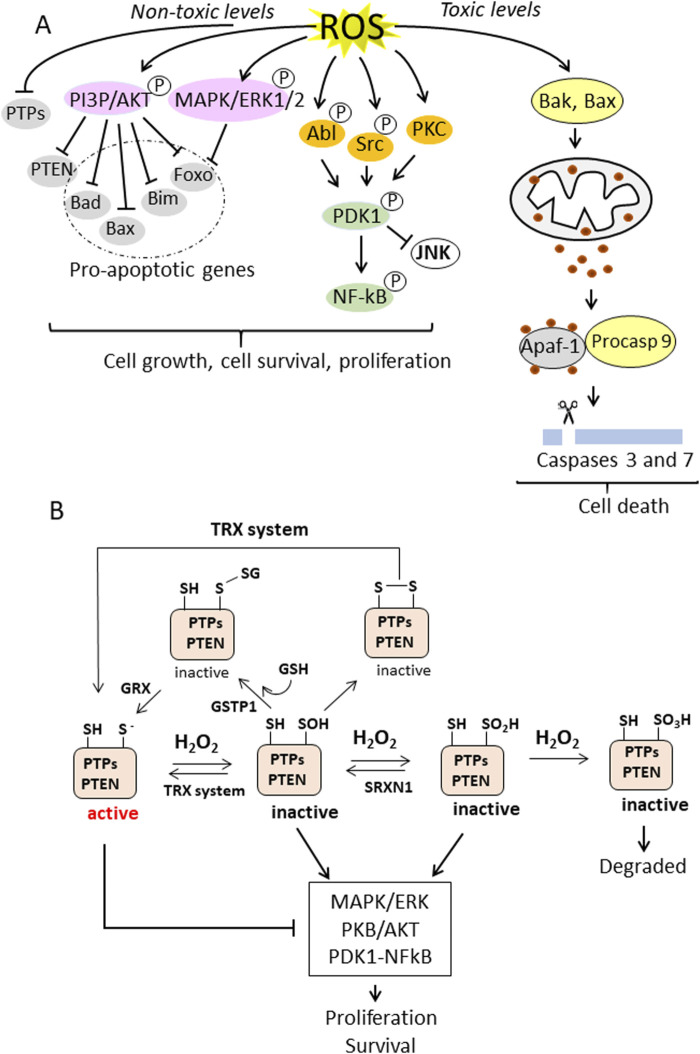
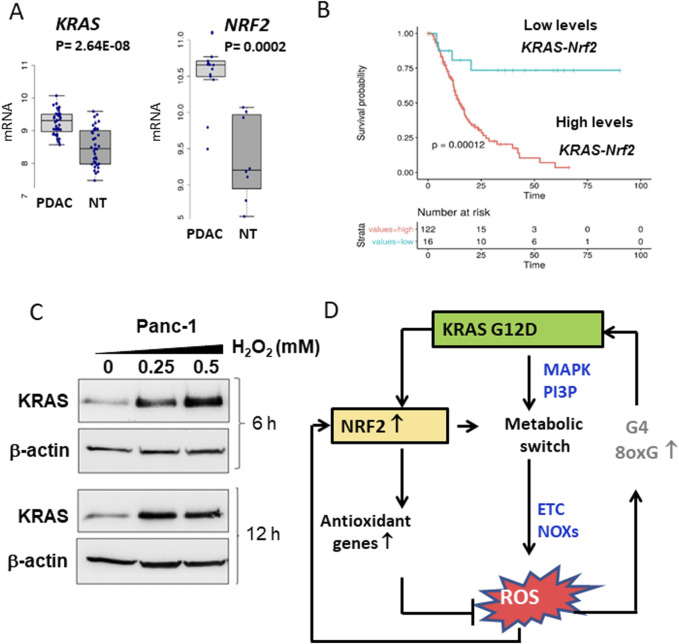
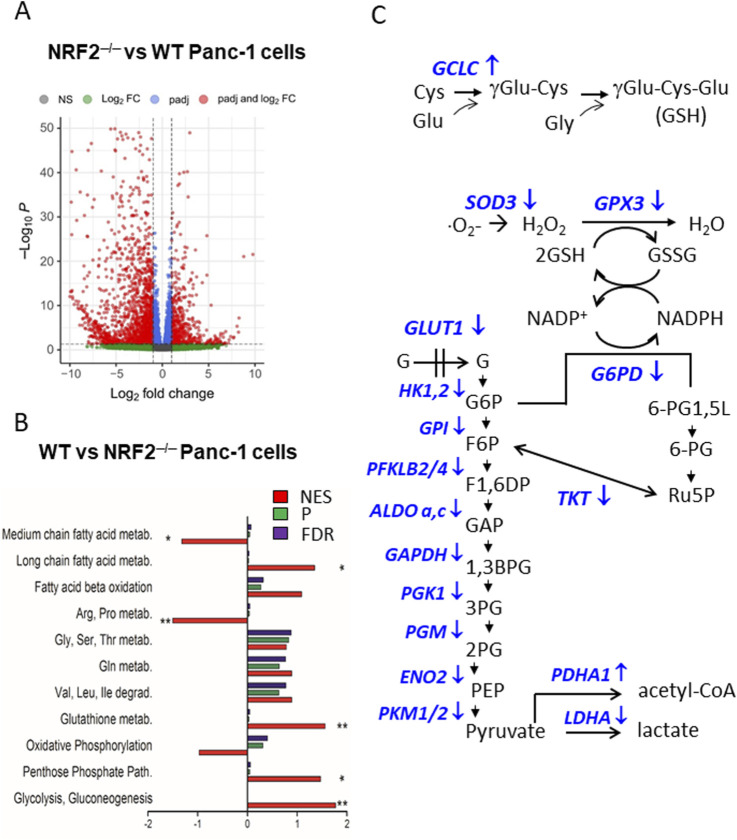
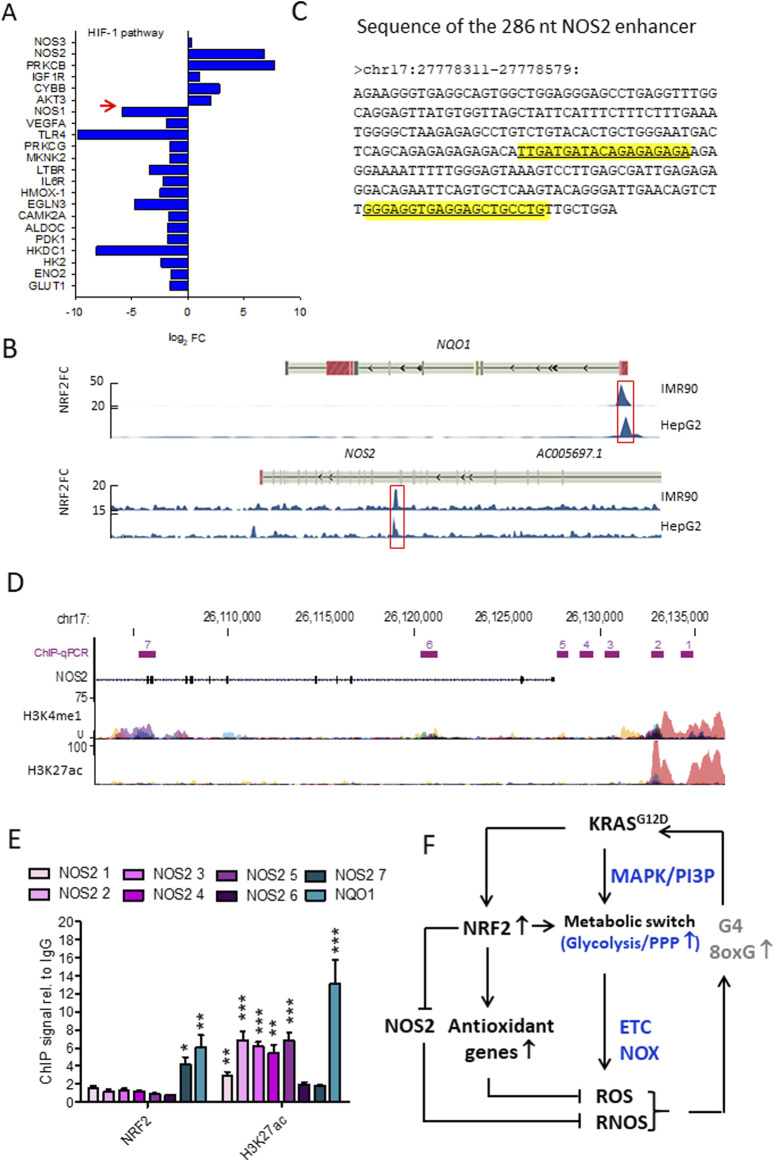
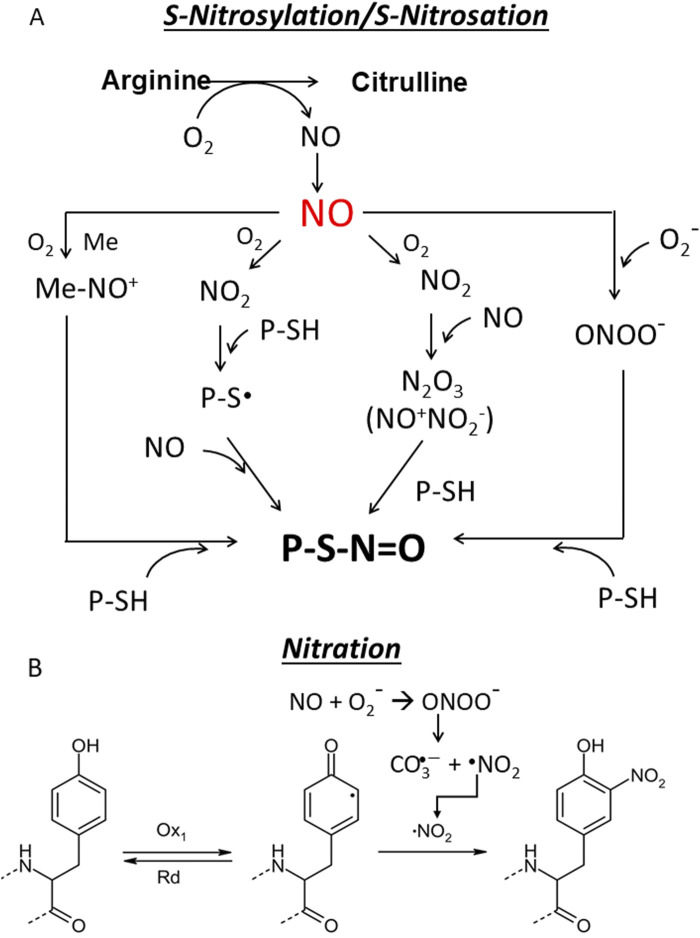
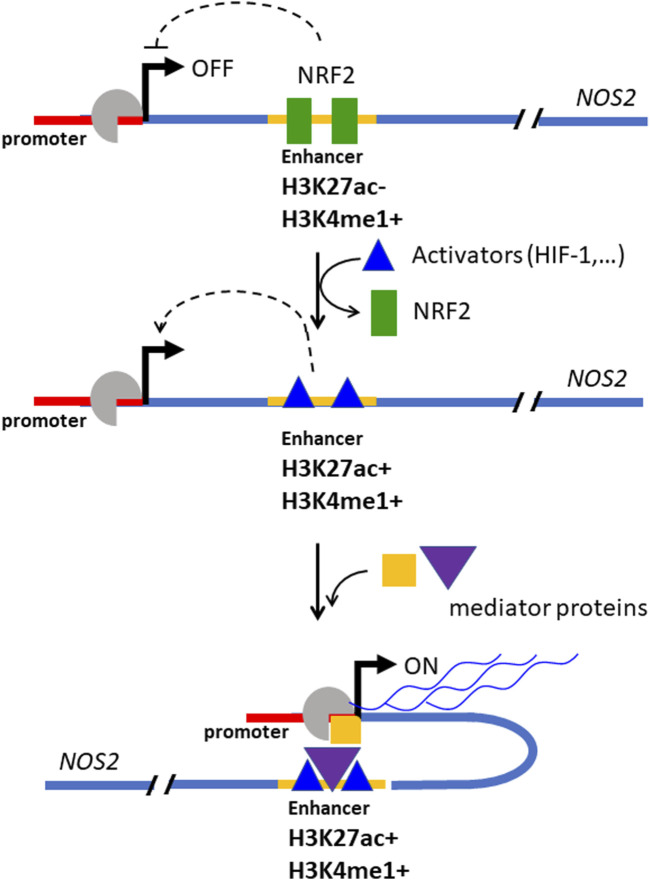
Cancer cells are subject to metabolic reprogramming, which leads to a sustained production of reactive oxygen species (ROS). Increased oxidative stress contributes to genomic instability and promotes malignant transformation. To counteract excessive ROS levels, cells activate nuclear factor erythroid 2-related factor 2 (NRF2), a key regulator of redox homeostasis that coordinates the transcription of a wide range of antioxidant and cytoprotective genes. This review examines the metabolic adaptations controlled by the KRAS-NRF2 axis under oxidative stress conditions. In addition, we highlight a novel function of NRF2 in regulating the expression of NOS2 by binding to a DNA enhancer element, thereby modulating the production of reactive nitrogen species (RNS). Finally, we discuss novel molecular strategies aimed at disrupting adaptive antioxidant responses in cancer cells and provide insights into combinatorial therapeutic approaches targeting redox balance in cancer.
Keywords: KRAS; NOS2; NRF2; PDAC; RNS; ROS; metabolic reprogramming.
Copyright © 2025 Rapozzi, Comuzzi, Di Giorgio and Xodo.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous