

This is a preprint.
An optogenetic toolkit for robust activation of FGF, BMP, & Nodal signaling in zebrafish
- PMID: 40568079
- PMCID: PMC12190770
- DOI: 10.1101/2025.04.17.649426
An optogenetic toolkit for robust activation of FGF, BMP, & Nodal signaling in zebrafish
Abstract
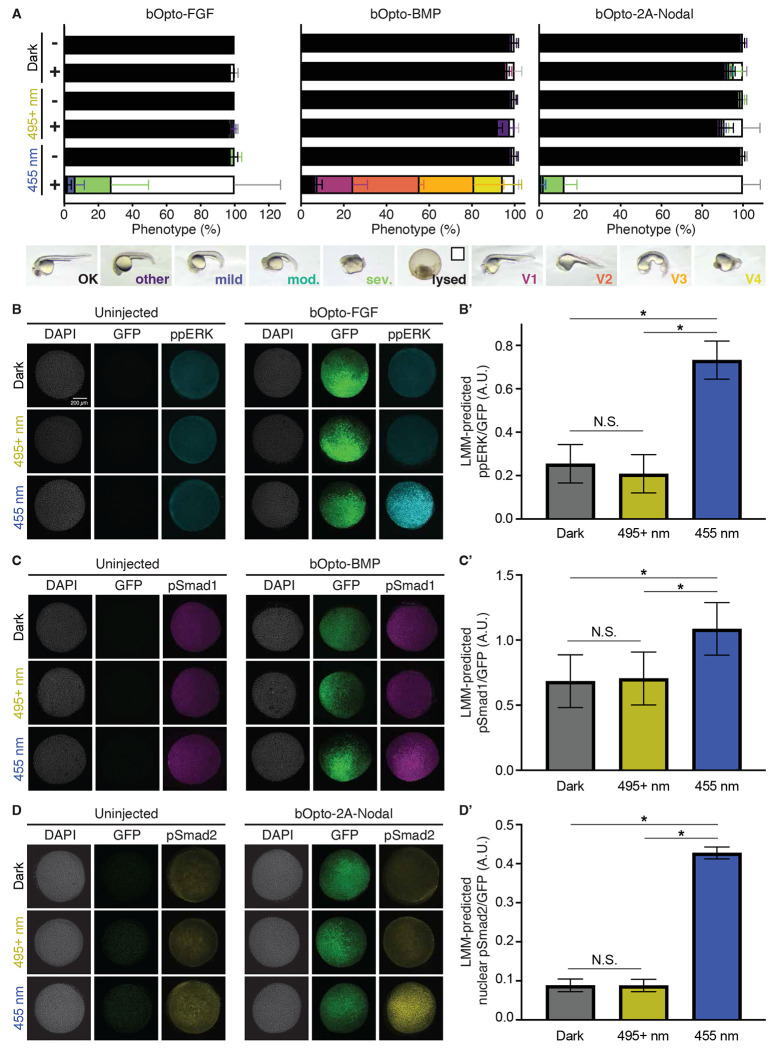
Cell signaling regulates a wide range of biological processes including development, homeostasis, and disease. Accessible technologies to precisely manipulate signaling have important applications in basic and translational research. Here, we introduce an optogenetic toolkit comprised of 1) a zebrafish-optimized FGF signaling activator, 2) a single-transcript Nodal signaling activator, and 3) a previously established BMP signaling activator. We thoroughly characterize this suite of tools in zebrafish embryos and show that they provide tunable, light-dependent spatiotemporal control of signaling in vivo. In response to blue light (~455 nm), receptor kinase domains fused to blue light-dimerizing LOV domains enable robust signaling activation with minimal ectopic activity in the dark or at wavelengths over 495 nm. Optogenetic activation by each tool is pathway-specific and results in increased expression of known target genes. Signaling is activated with rapid on/off kinetics, and activation strength depends on light irradiance. Finally, we demonstrate spatially localized signaling activation with our optimized FGF activator. Together, our results establish this optogenetic toolkit as a potent experimental platform to rapidly, directly, and adjustably activate FGF, BMP, and Nodal signaling in zebrafish embryos.
Keywords: BMP; FGF; Nodal; embryogenesis; molecular optogenetics; signaling; zebrafish.
Conflict of interest statement
Competing interests: No competing interests declared.
Figures






References
-
- Belacortu Y. and Paricio N. (2011). Drosophila as a model of wound healing and tissue regeneration in vertebrates. Dev Dyn 240, 2379–2404. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials