

Microbial cross-feeding stabilized by segregation of a dependent mutant from its independent ancestor
- PMID: 40568968
- PMCID: PMC12279053
- DOI: 10.1093/ismejo/wraf131
Microbial cross-feeding stabilized by segregation of a dependent mutant from its independent ancestor
Abstract
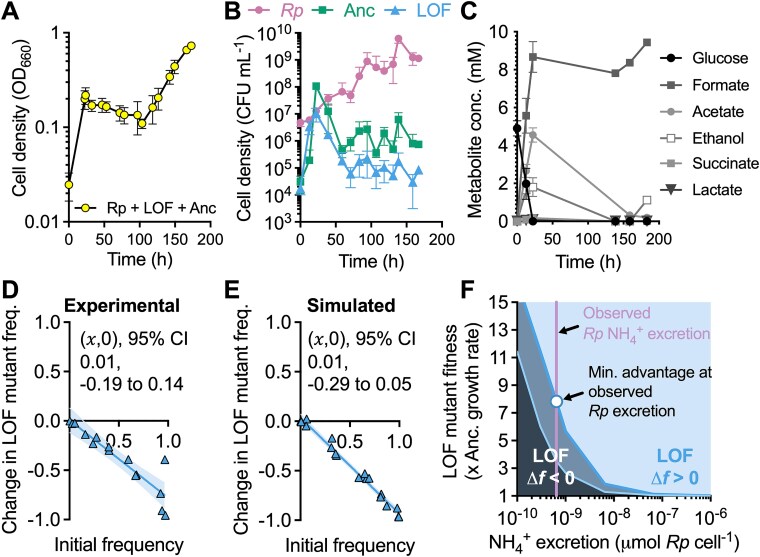
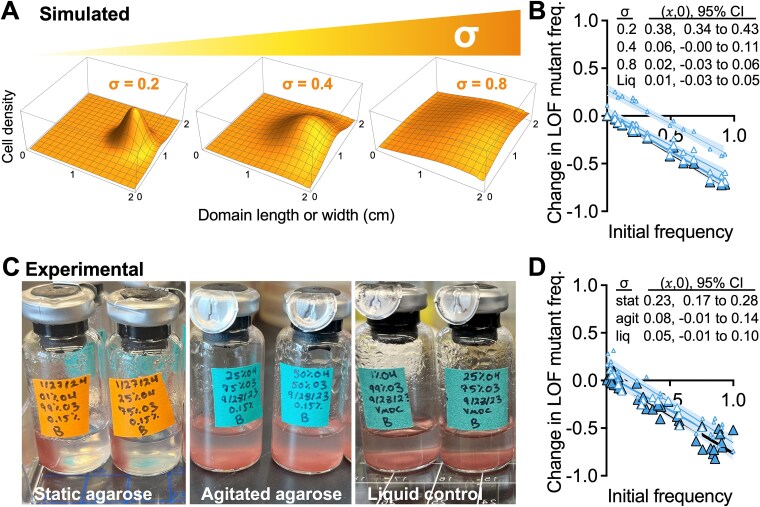
Microbial gene loss is hypothesized to be beneficial when gene function is costly, and the gene product can be replaced via cross-feeding from a neighbor. However, cross-fed metabolites are often only available at low concentrations, limiting the growth rates of gene-loss mutants that are dependent on those metabolites. Here we define conditions that support a loss of function mutant in a three-member bacterial community of (i) N2-utilizing Rhodopseudomonas palustris as an NH4+-excreting producer, (ii) N2-utilizing Vibrio natriegens as the ancestor, and (iii) a V. natriegens N2-utilizaton mutant that is dependent on the producer for NH4+. Using experimental and simulated cocultures, we found that the ancestor outcompeted the mutant due to low NH4+ availability under uniform conditions where both V. natriegens strains had equal access to nutrients. However, spatial structuring that increasingly segregated the mutant from the ancestor, while maintaining access to NH4+ from the producer, allowed the mutant to avoid extinction. Counter to predictions, mutant enrichment under spatially structured conditions did not require a growth rate advantage from gene loss and the mutant coexisted with its ancestor. Thus, cross-feeding can originate from loss-of-function mutations that are otherwise detrimental, provided that the mutant can segregate from a competitive ancestor.
Keywords: Rhodopseudomonas palustris; Vibrio natriegens; Black Queen Hypothesis; cross-feeding; excretion; gene loss; microbial interactions; microbial physiology; mutualism; syntrophy.
© The Author(s) 2025. Published by Oxford University Press on behalf of the International Society for Microbial Ecology.
Conflict of interest statement
None declared.
Figures







References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
