

C1orf50 Drives Malignant Melanoma Progression Through the Regulation of Stemness
- PMID: 40571314
- PMCID: PMC12216575
- DOI: 10.21873/cgp.20518
C1orf50 Drives Malignant Melanoma Progression Through the Regulation of Stemness
Abstract
Background/aim: Recent advancements in omics analysis have significantly enhanced our understanding of the molecular pathology of malignant melanoma, leading to the development of novel therapeutic strategies that target specific vulnerabilities within the disease. Despite these improvements, the factors contributing to the poor prognosis of patients with malignant melanoma remain incompletely understood. The aim of this study was to investigate the role of C1orf50 (Chromosome 1 open reading frame 50), a gene previously of unknown function, as a prognostic biomarker in melanoma.
Materials and methods: We performed comprehensive transcriptome data analysis and subsequent functional validation of the human Skin Cutaneous Melanoma project from The Cancer Genome Atlas (TCGA).
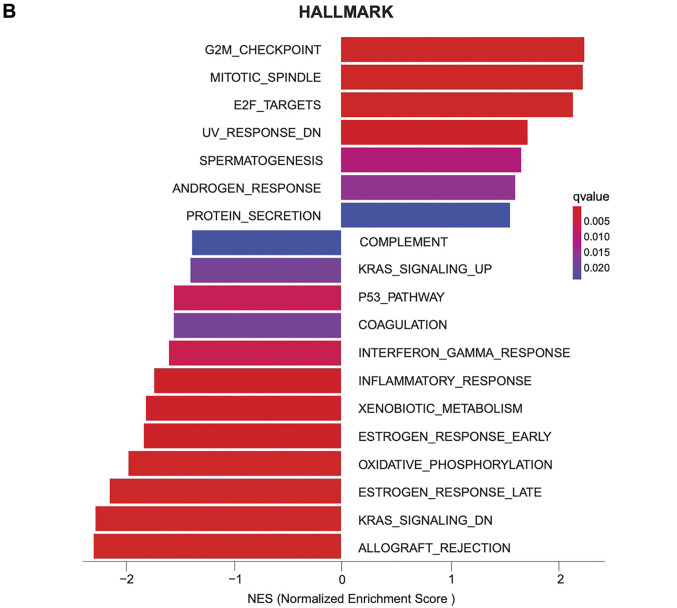
Results: Elevated expression levels of C1orf50 correlated with worse survival outcomes. Mechanistically, we revealed that C1orf50 plays a significant role in the regulation of cell cycle processes and cancer cell stemness, providing a potential avenue for novel therapeutic interventions in melanoma.
Conclusion: This study is the first to identify C1orf50 as a prognostic biomarker in melanoma. The clinical relevance of our results sheds light on the importance of further investigation into the biological mechanisms underpinning C1orf50's impact on melanoma progression and patient prognosis.
Keywords: C1orf50; YAP/TAZ; cancer stem cells; melanoma.
Copyright © 2025, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved.
Conflict of interest statement
MHR is a member of Universal DX’s Scientific Advisory Board. However, this company had no influence on support, design, execution, data analysis, or other aspects of this study.
Figures









References
-
- Krug K, Jaehnig EJ, Satpathy S, Blumenberg L, Karpova A, Anurag M, Miles G, Mertins P, Geffen Y, Tang LC, Heiman DI, Cao S, Maruvka YE, Lei JT, Huang C, Kothadia RB, Colaprico A, Birger C, Wang J, Dou Y, Wen B, Shi Z, Liao Y, Wiznerowicz M, Wyczalkowski MA, Chen XS, Kennedy JJ, Paulovich AG, Thiagarajan M, Kinsinger CR, Hiltke T, Boja ES, Mesri M, Robles AI, Rodriguez H, Westbrook TF, Ding L, Getz G, Clauser KR, Fenyö D, Ruggles KV, Zhang B, Mani DR, Carr SA, Ellis MJ, Gillette MA, Clinical Proteomic Tumor Analysis Consortium Proteogenomic landscape of breast cancer tumorigenesis and targeted therapy. Cell. 2020;183(5):1436–1456.e31. doi: 10.1016/j.cell.2020.10.036. - DOI - PMC - PubMed
-
- Tanaka A, Ogawa M, Zhou Y, Namba K, Hendrickson RC, Miele MM, Li Z, Klimstra DS, Buckley PG, Gulcher J, Wang JY, Roehrl MHA. Proteogenomic characterization of primary colorectal cancer and metastatic progression identifies proteome-based subtypes and signatures. Cell Rep. 2024;43(2):113810. doi: 10.1016/j.celrep.2024.113810. - DOI - PMC - PubMed
-
- Tanaka A, Ogawa M, Zhou Y, Otani Y, Hendrickson RC, Miele MM, Li Z, Klimstra DS, Wang JY, Roehrl MH. Proteogenomic characterization of pancreatic neuroendocrine tumors uncovers hypoxia and immune signatures in clinically aggressive subtypes. iScience. 2024;27(8):110544. doi: 10.1016/j.isci.2024.110544. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical