

Extrahepatic Replication and Genomic Signatures of the Hepatitis E Virus in the Kidney
- PMID: 40575997
- PMCID: PMC12203452
- DOI: 10.1111/liv.70183
Extrahepatic Replication and Genomic Signatures of the Hepatitis E Virus in the Kidney
Abstract
Introduction: The hepatitis E virus (HEV; species Paslahepevirus balayani) is a common human pathogenic and zoonotic virus that can cause both acute fulminant and chronic hepatitis. Despite its reputation as a hepatotropic virus, HEV infection is also associated with a number of extrahepatic diseases, including kidney disorders. However, the extent to which HEV replicates in kidney cells remains unclear. The present study aims to investigate the capacity of HEV to propagate in kidney cells in vitro and to assess whether HEV displays mutational signatures that correlate with compartmentalisation in vivo.
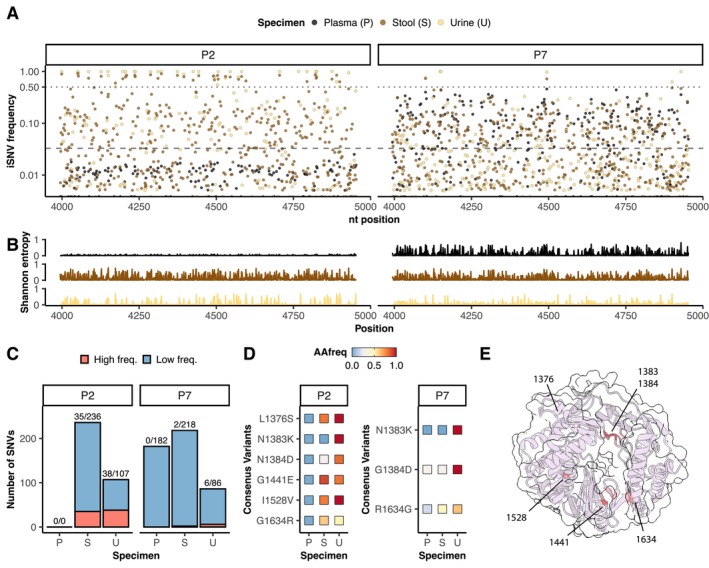
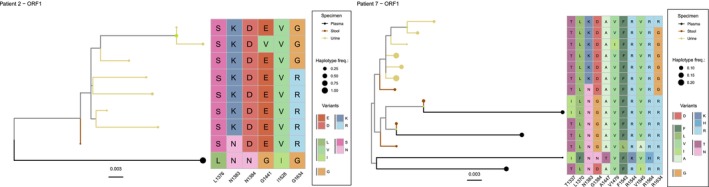
Methods: We use HEV cell culture models to study the replication cycle and the effect of antivirals in human kidney cell lines and primary cells. In addition, we identified patients with chronic HEV infection (n = 9) from which we then sequenced the viral RNA of urine, stools and plasma to analyse the viral sequence composition, to assess intra-host diversity and compartmentalisation (n = 2).
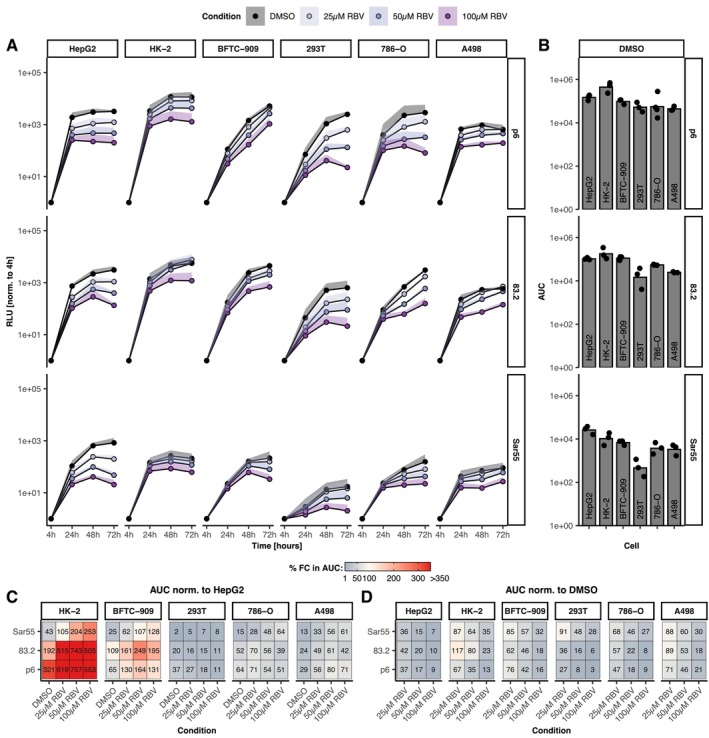
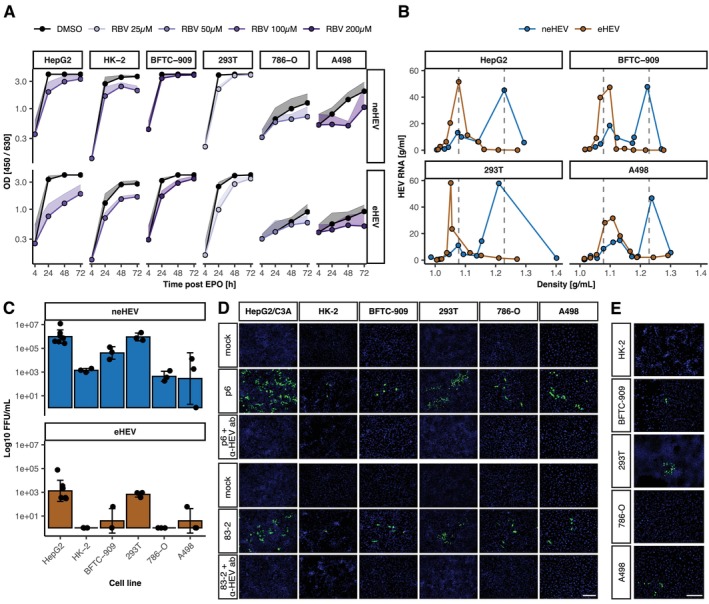
Results: A wide range of human kidney cell lines as well as primary cells supports viral entry, replication and propagation of HEV in vitro. Interestingly, the broad-spectrum antiviral ribavirin was less effective in inhibiting HEV replication in some kidney cells. Sequencing of HEV RNA-directed RNA polymerase coding region from plasma, stool and urine and subsequent phylogenetic analysis revealed diversification of HEV into tissue-specific viral subpopulations. In particular, the viruses derived from urine were found to be distinct from those derived from plasma and stool.
Conclusions: In conclusion, kidney cells support the propagation of HEV in vitro and exhibit reduced sensitivity to antiviral treatment. Furthermore, HEV patient-derived sequences demonstrated compartmentalisation into distinct clusters that correlated with sample source. Collectively, these data indicate the potential for extrahepatic replication of HEV, which may result in clinically significant disease or serve as a reservoir for patient relapse.
Trial registration: HepNet-SofE study (NCT03282474).
Keywords: compartmentalization; hepatitis E; kidney impairment.
© 2025 The Author(s). Liver International published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- World Health Organization , “Hepatitis E Fact Sheets [Internet],” (2022) cited 2023 Jan 3, https://www.who.int/news‐room/fact‐sheets/detail/hepatitis‐e.
-
- Dalton H. R., Kamar N., Baylis S. A., Moradpour D., Wedemeyer H., and Negro F., “EASL Clinical Practice Guidelines on Hepatitis E Virus Infection,” Journal of Hepatology 68, no. 6 (2018): 1256–1271. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
