

Gut microbial metabolite butyrate suppresses hepatocellular carcinoma growth via CXCL11-dependent enhancement of natural killer cell infiltration
- PMID: 40576244
- PMCID: PMC12218501
- DOI: 10.1080/19490976.2025.2519706
Gut microbial metabolite butyrate suppresses hepatocellular carcinoma growth via CXCL11-dependent enhancement of natural killer cell infiltration
Abstract
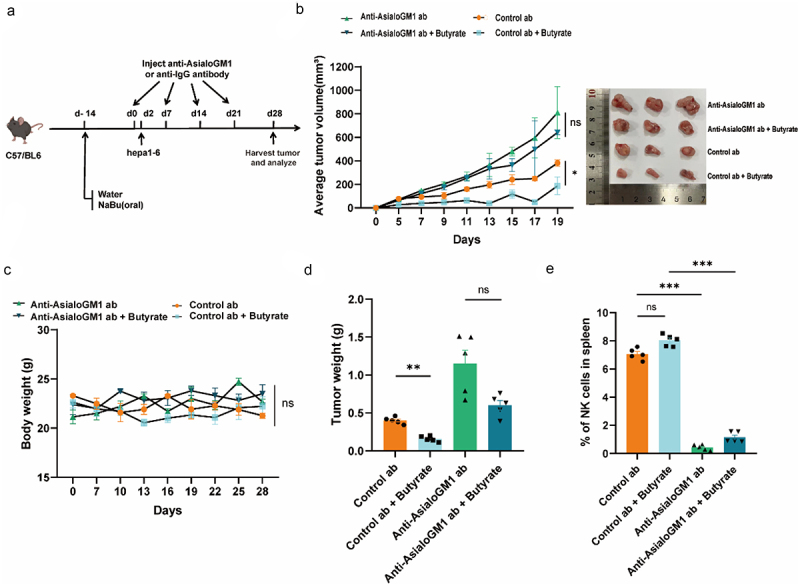
Gut microbiota-derived butyrate plays a vital role in attenuating hepatocellular carcinoma (HCC) in murine models. However, the precise molecular mechanisms by which butyrate exerts its effects are largely undefined. Plasma short-chain fatty acids (SCFAs) were quantitatively measured by using gas chromatography-mass spectrometry (GC-MS) to access their association with HCC prognosis. Tumor-infiltrating immune cells were characterized by flow cytometry. The interactions between butyrate and natural killer (NK) cells were studied using in vitro assays, including migration, cytotoxic degranulation, and co-culture experiments. In vivo validation was conducted through neutralization experiments. The molecular pathways regulated by butyrate were further investigated by employing RNA sequencing (RNA-seq), chromatin immunoprecipitation sequencing (ChIP-seq), and Assay for Transposase-Accessible Chromatin using sequencing (ATAC-seq). A positive correlation was observed between elevated plasma butyrate levels and improved prognosis in HCC patients. Notably, butyrate inhibited tumor progression by enhancing NK cell infiltration into tumor tissues. Mechanistically, butyrate stimulated cytokine secretion, notably significantly enhancing the production of the chemokine CXCL11, thereby facilitating NK cell infiltration. Gene Set Enrichment Analysis (GSEA) of hepatic tumor cell lines revealed that the chemokine signaling and NK cell-mediated cytotoxicity pathways were upregulated following butyrate stimulation. Furthermore, transcriptomic and epigenomic analyses showed that exposure to butyrate induced de novo chromatin accessibility and enhancer remodeling, regulated by STAT family transcription factors. Our study demonstrated that butyrate was able to enhance the expression of CXCL11. This is likely attributed to chromatin remodeling, and then promoting NK cell infiltration and exerting effective anti-tumor effects on HCC.
Keywords: Gut microbiota; butyrate; epigenetic; hepatocellular carcinoma; natural killer cells.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures






References
-
- Waked I, Alsammany S, Tirmazy SH, Rasul K, Bani-Issa J, Abdel-Razek W, Omar A, Shafik A, Eid S, Abdelaal A, et al. Multidisciplinary consensus recommendations for management of hepatocellular carcinoma in Middle East and North Africa region. Liver Int. 2023;43(10):2062–16. doi: 10.1111/liv.15685. - DOI - PubMed
-
- Song Q, Zhang X, Liu W, Wei H, Liang W, Zhou Y, Ding Y, Ji F, ho-Kwan Cheung A, Wong N, et al. Bifidobacterium pseudolongum-generated acetate suppresses non-alcoholic fatty liver disease-associated hepatocellular carcinoma. J Hepatol. 2023;79(6):1352–1365. doi: 10.1016/j.jhep.2023.07.005. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous