

A root-specific NLR network mediates immune signaling of resistance genes against plant parasitic nematodes
- PMID: 40581623
- PMCID: PMC12236159
- DOI: 10.1093/plcell/koaf145
A root-specific NLR network mediates immune signaling of resistance genes against plant parasitic nematodes
Abstract
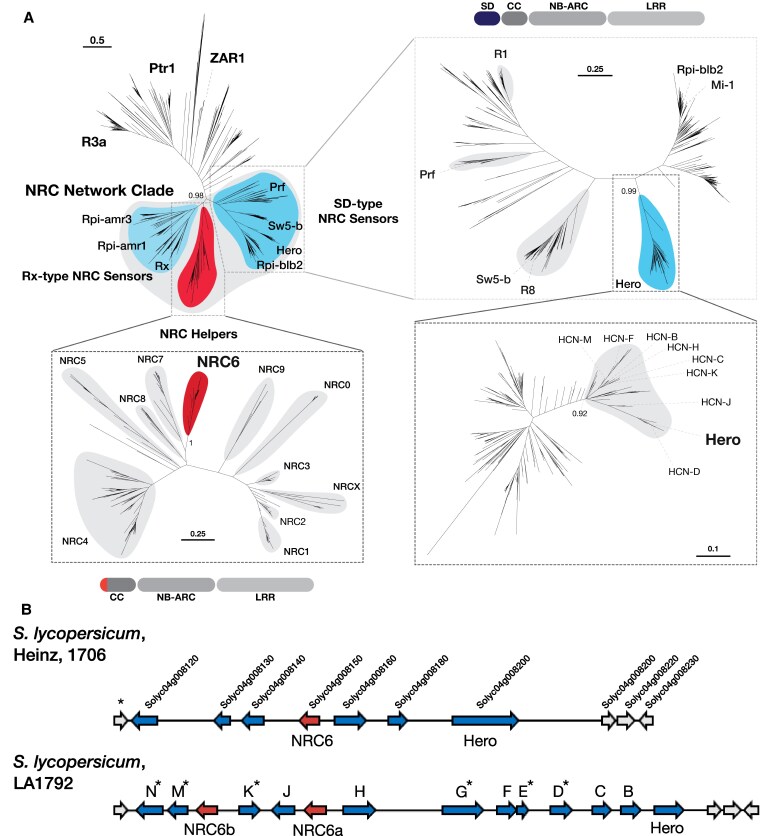
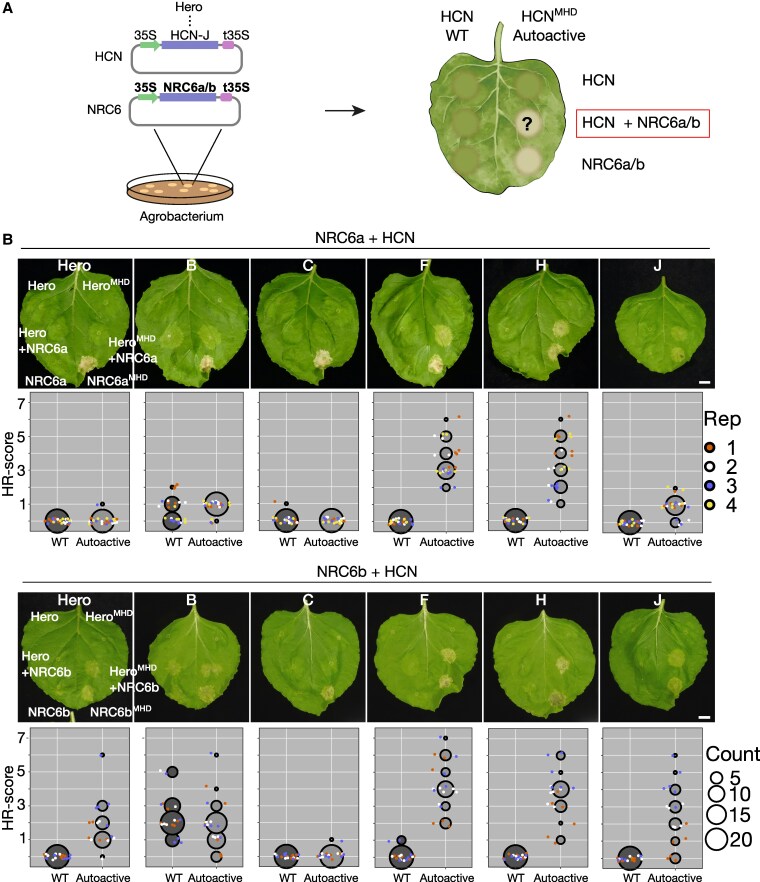
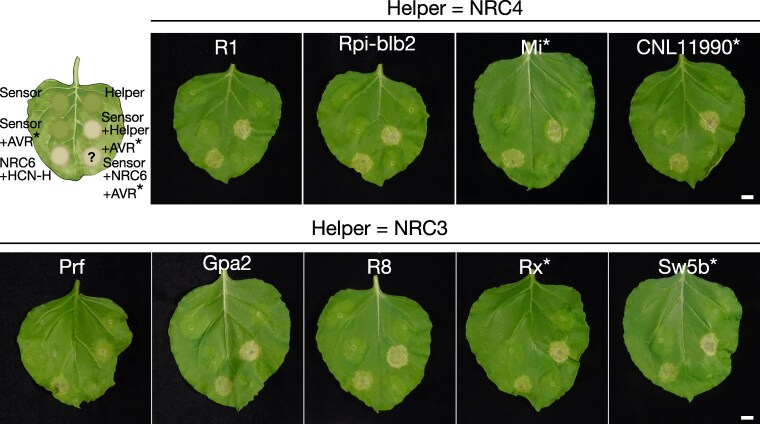
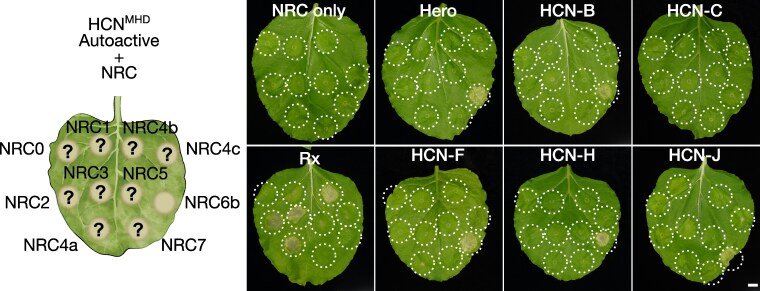
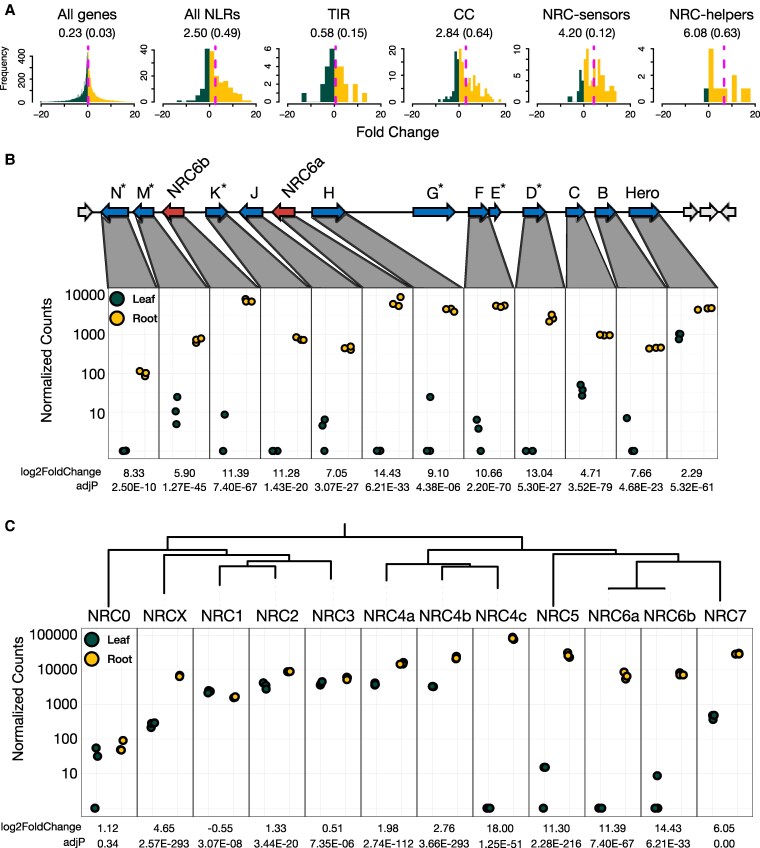
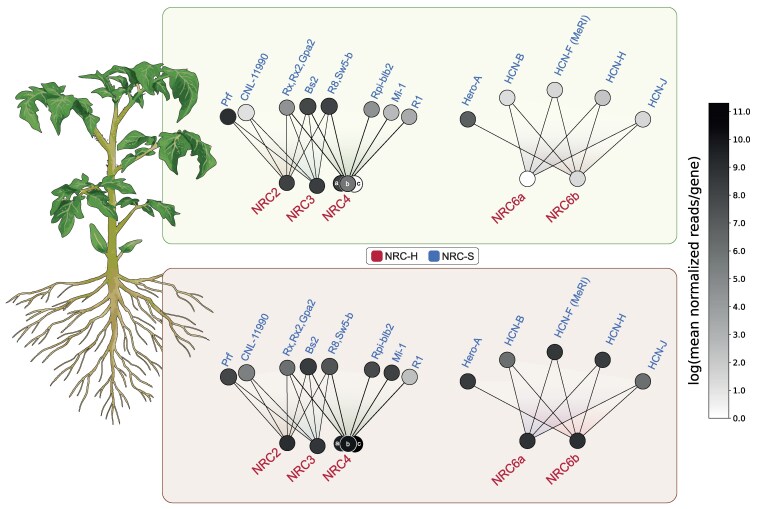
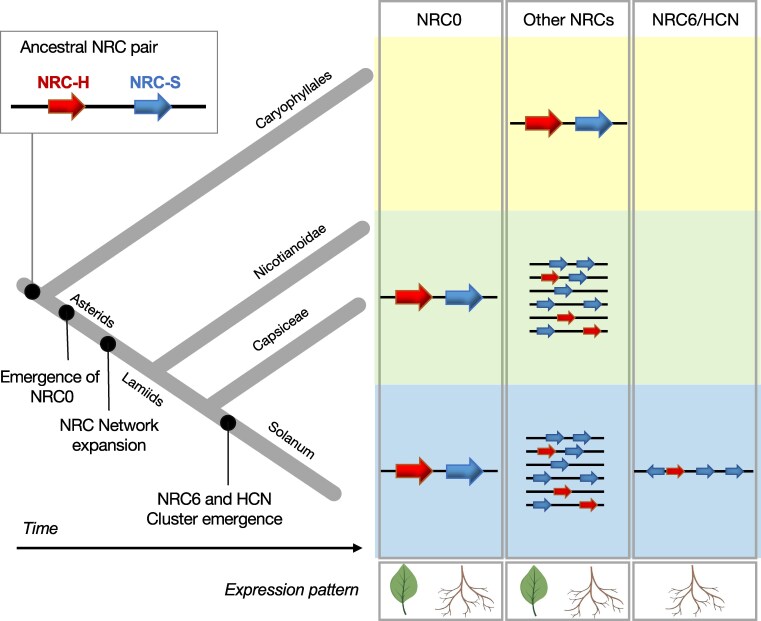
Plant nucleotide-binding domain and leucine-rich repeat immune receptors (NLRs) confer disease resistance to many foliar and root parasites. However, the extent to which NLR-mediated immunity is differentially regulated between plant organs is poorly known. Here, we show that a large cluster of tomato (Solanum lycopersicum) genes, encoding the cyst and root-knot nematode disease resistance proteins Hero and MeR1 as well as the NLR helper NLR required for cell death 6 (NRC6), is nearly exclusively expressed in the roots. This root-specific gene cluster emerged in Solanum species about 21 million years ago through gene duplication of the ancient asterid NRC network. NLR sensors in this gene cluster function exclusively through NRC6 helpers to trigger hypersensitive cell death. These findings indicate that the NRC6 gene cluster has sub-functionalized from the larger NRC network to specialize in mediating resistance against root pathogens, including cyst and root-knot nematodes. We propose that some NLR gene clusters and networks may have evolved organ-specific gene expression as an adaptation to particular parasites and to reduce the risk of autoimmunity.
© The Author(s) 2025. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Conflict of interest statement
Conflict of interest statement. S.K. and J.K. have filed patents on NLR biology and receive funding from industry on NLR biology. S.K. is a co-founder of start-up companies that focus on plant disease resistance. K.E., M.G., R.F., and A.V. have filed patents on nematode resistance genes.
Figures


Similar articles
-
The SlDOF9-SlSWEET17 Module: a Switch for Controlling Sugar Distribution Between Nematode Induced Galls and Roots in Tomato.Adv Sci (Weinh). 2025 Jul;12(28):e2501771. doi: 10.1002/advs.202501771. Epub 2025 May 11. Adv Sci (Weinh). 2025. PMID: 40349184 Free PMC article.
-
NRC Immune receptor networks show diversified hierarchical genetic architecture across plant lineages.Plant Cell. 2024 Sep 3;36(9):3399-3418. doi: 10.1093/plcell/koae179. Plant Cell. 2024. PMID: 38922300 Free PMC article.
-
Stacking potato NLR genes activates a calcium-dependent protein kinase and confers broad-spectrum disease resistance to late blight.J Integr Plant Biol. 2025 Jul;67(7):1910-1927. doi: 10.1111/jipb.13892. Epub 2025 Mar 24. J Integr Plant Biol. 2025. PMID: 40125812
-
Pattern Recognition Receptors in Plant Immunity.Adv Exp Med Biol. 2025;1476:425-451. doi: 10.1007/978-3-031-85340-1_17. Adv Exp Med Biol. 2025. PMID: 40622553 Review.
-
Natural Resources Resistance to Tomato Spotted Wilt Virus (TSWV) in Tomato (Solanum lycopersicum).Int J Mol Sci. 2021 Oct 12;22(20):10978. doi: 10.3390/ijms222010978. Int J Mol Sci. 2021. PMID: 34681638 Free PMC article. Review.
References
-
- Ahn H-K, Lin X, Olave-Achury AC, Derevnina L, Contreras MP, Kourelis J, Wu C-H, Kamoun S, Jones JDG. Effector-dependent activation and oligomerization of plant NRC class helper NLRs by sensor NLR immune receptors Rpi-amr3 and Rpi-amr1. EMBO J. 2023:42(5):e111484. 10.15252/embj.2022111484 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- Gatsby Charitable Foundation
- BB/WW002221/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/V002937/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/000PR9795/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/000PR9796/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BASF Plant Science/ERC_/European Research Council/International
- Japan Science and Technology Agency
- JPMJPR21D1/Precursory Research for Embryonic Science and Technology
- NSTC 112-2628-B-001-007/2030 Cross-Generation Young Scholars Program of the National Science and Technology Council
- AS-GC-111-L02/Academia Sinica Grand Challenge Program
- Ga470/1/1-3/DFG
- 464864389/DFG Walter Benjamin Programme
LinkOut - more resources
Full Text Sources
