

Alzheimer's disease brain-derived tau extracts show differential processing and transcriptional effects in human astrocytes
- PMID: 40585369
- PMCID: PMC12205603
- DOI: 10.1016/j.isci.2025.112793
Alzheimer's disease brain-derived tau extracts show differential processing and transcriptional effects in human astrocytes
Abstract
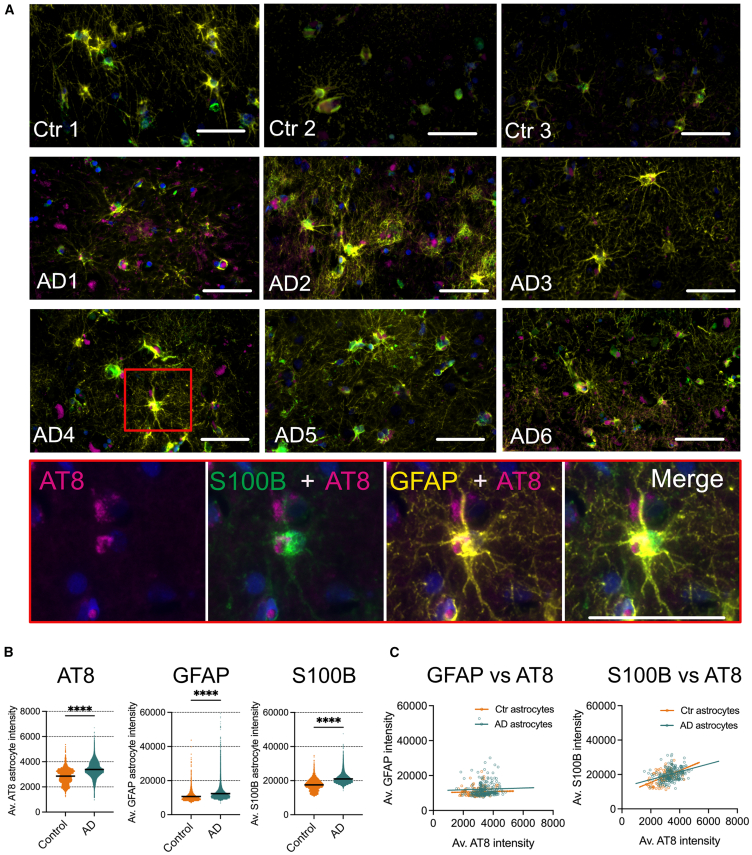
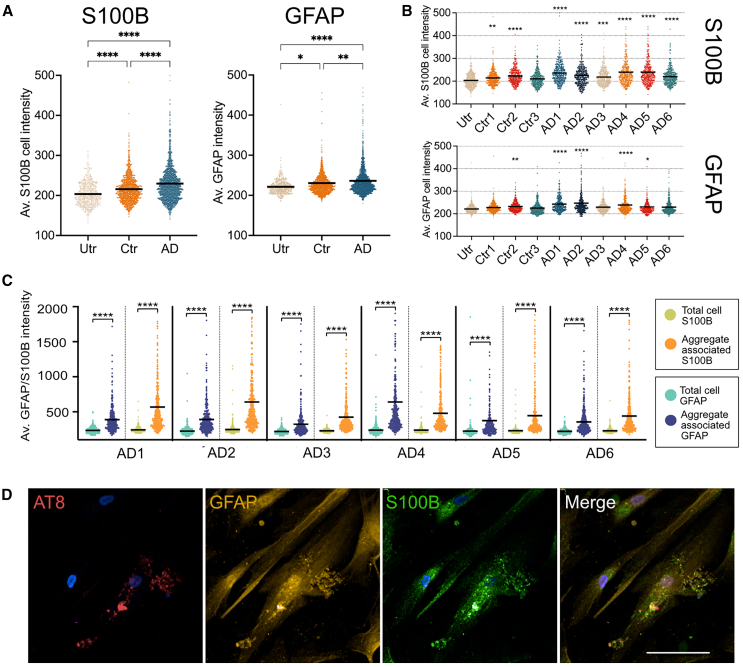
Post-translational modifications of tau, including phosphorylation at specific residues, are closely linked with tau seeding ability and clinical disease progression. While most previous evidence has focused on neuronal tau spread, evidence supports a similar role for astrocytes. Here, we demonstrate that well characterized tau aggregates isolated from postmortem Alzheimer's disease brain are internalized and processed by control human-induced pluripotent stem cell-derived astrocytes. Differences in the efficiency of tau internalization, clearance and/or seeding were noted, which reflect molecular properties of tau and/or co-factors in brain extracts. We observed a direct relationship between tau handling by astrocytes and astrocyte transcriptomic changes. Dysregulated genes include several previously identified as upregulated in reactive astrocytes in Alzheimer's brain, as well as those implicated in pathological tau clearance by autophagy and other pathways. The study provides insights into the complex interplay between tau molecular diversity and astrocyte responses in Alzheimer's disease.
Keywords: Cell biology; Cellular neuroscience; Neuroscience; Omics.
© 2025 The Authors.
Conflict of interest statement
The author declare that they have no competing interests.
Figures





References
-
- Ferrari-Souza J.P., Ferreira P.C.L., Bellaver B., Tissot C., Wang Y.T., Leffa D.T., Brum W.S., Benedet A.L., Ashton N.J., De Bastiani M.A., et al. Astrocyte biomarker signatures of amyloid-beta and tau pathologies in Alzheimer's disease. Mol. Psychiatry. 2022;27:4781–4789. doi: 10.1038/s41380-022-01716-2. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
