

Resolving the APN controversy in PEDV infection: Comparative kinetic characterization through single-virus tracking
- PMID: 40587580
- PMCID: PMC12233901
- DOI: 10.1371/journal.ppat.1013317
Resolving the APN controversy in PEDV infection: Comparative kinetic characterization through single-virus tracking
Abstract
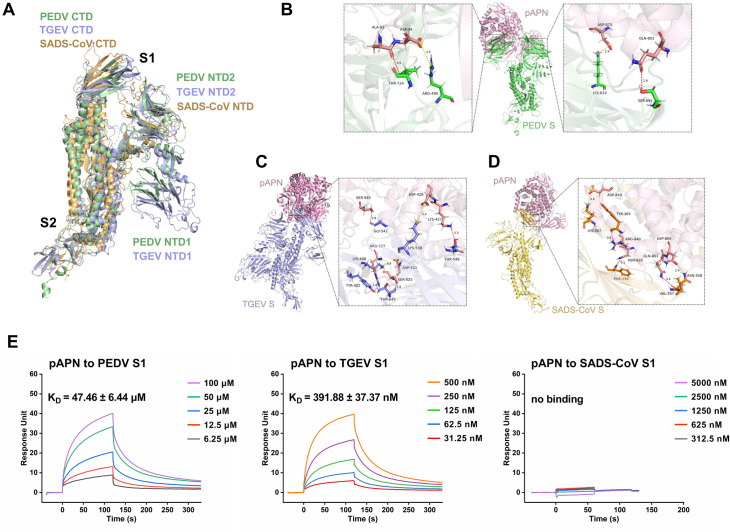
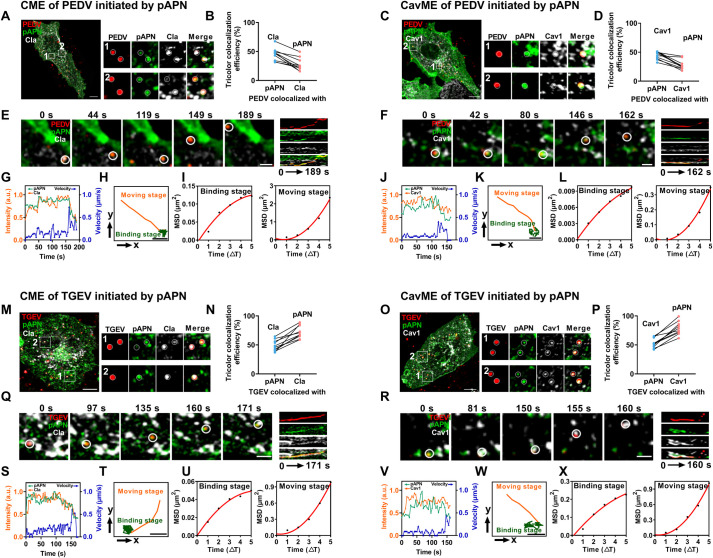
Aminopeptidase N (APN) plays multiple roles in various physiological processes, with its function as a viral receptor in several coronaviruses being one of the most prominent. However, the role of porcine APN (pAPN) in porcine epidemic diarrhea virus (PEDV) has remained controversial. Single-virus tracking enables a more comprehensive dynamic dissection of pAPN utilization during virus entry. In this study, a comparative analysis of pAPN usage by PEDV, transmissible gastroenteritis virus (TGEV), and swine acute diarrhea syndrome coronavirus (SADS-CoV) provides more precise and quantitative insights into pAPN's specific role in PEDV entry. Here, we used molecular docking and surface plasmon resonance (SPR) to demonstrate that pAPN binds to PEDV, with lower affinity than to TGEV. However, pAPN facilitates PEDV replication through internalization only in susceptible cells, not in non-susceptible cells. Using single-virus tracking, we observed that pAPN triggers PEDV internalization via clathrin- and caveolae-mediated endocytosis, resembling a receptor-mediated process. pAPN participates in 35% of PEDV internalization events, but mediates 80% of TGEV internalization, with pAPN-mediated PEDV internalization occurring approximately 60 s slower than TGEV. The dynamic differences in the internalization of PEDV and TGEV mediated by pAPN primarily arise during the binding stage prior to the initiation of accelerated directional movement, whereas their durations of movement are comparable. Additionally, we found that the internalization dynamics of porcine deltacoronavirus (PDCoV), which also uses pAPN as a receptor, are similar to those of TGEV. These findings resolve the controversy surrounding pAPN's role in PEDV entry, and highlight the dynamic differences in PEDV, TGEV, PDCoV, and SADS-CoV internalization via pAPN at single-virus level, providing a novel theoretical basis for the potential receptor evaluation from kinetic perspective, which could significantly contribute to the development of strategies against future PEDV outbreaks.
Copyright: © 2025 An et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Miscellaneous
