

Several groups of LysM-RLKs are involved in symbiotic signal perception and arbuscular mycorrhiza establishment
- PMID: 40593575
- PMCID: PMC12217622
- DOI: 10.1038/s41467-025-60717-1
Several groups of LysM-RLKs are involved in symbiotic signal perception and arbuscular mycorrhiza establishment
Abstract
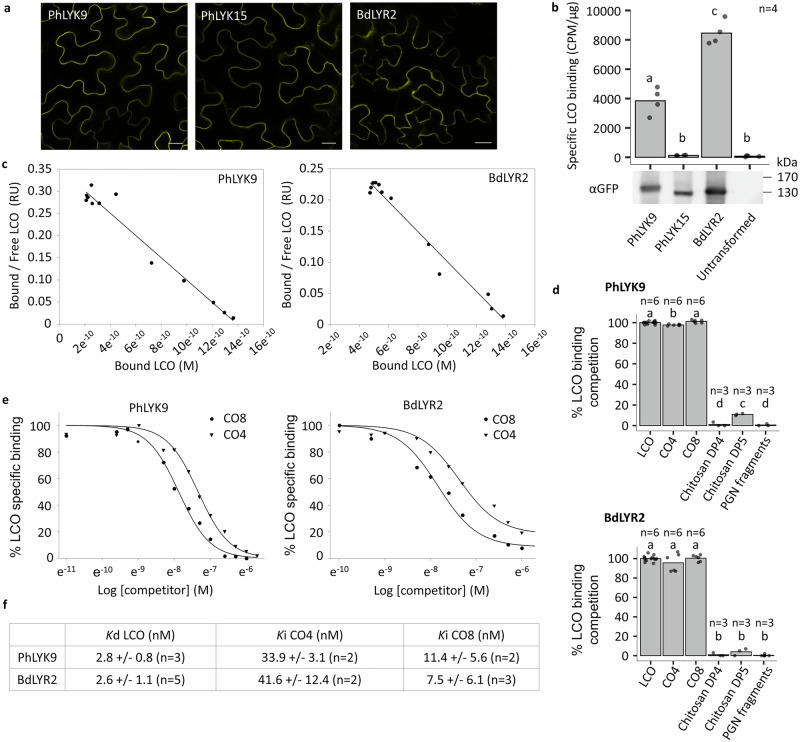
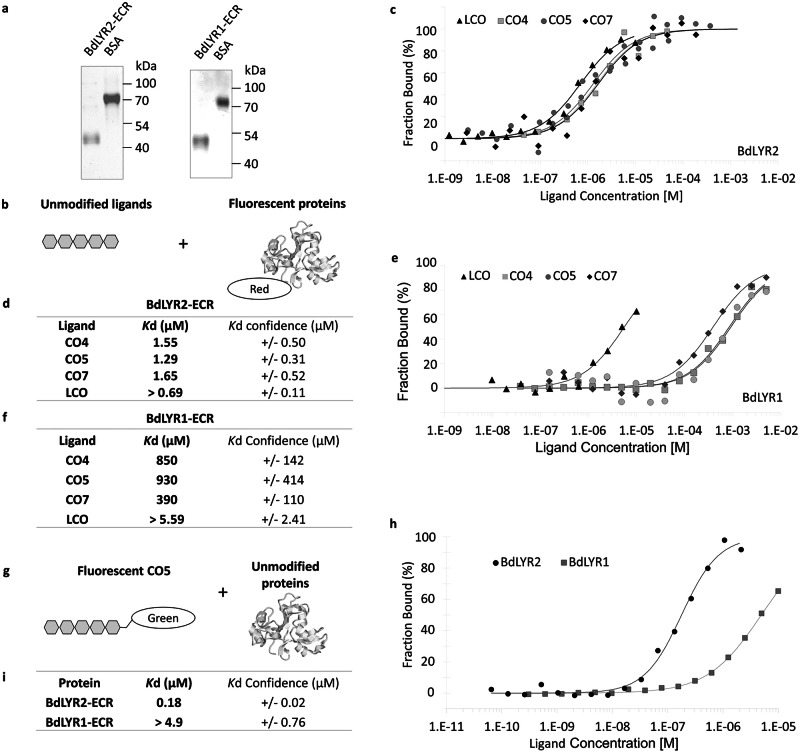
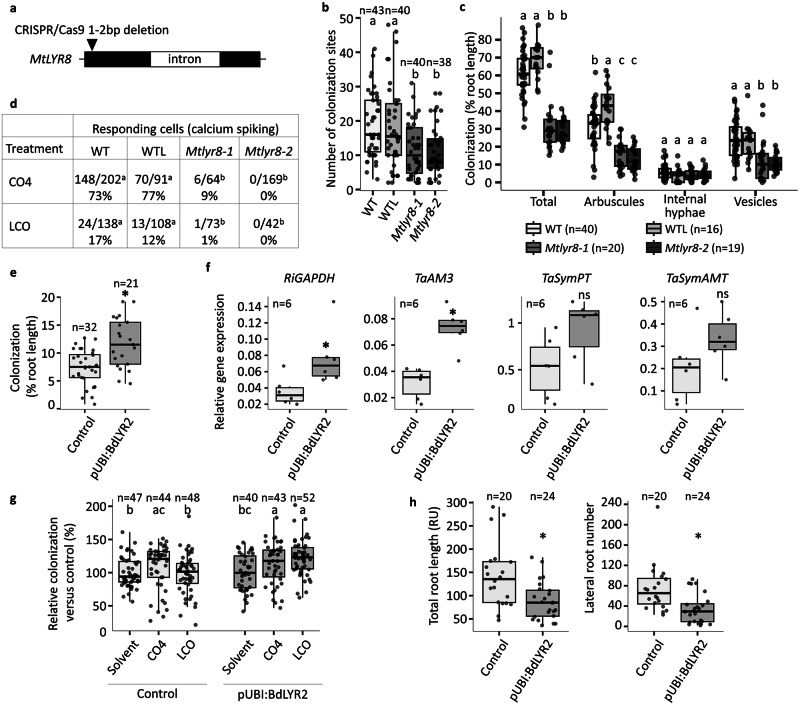
Lipo-chitooligosaccharides (LCO) and short-chain chitooligosaccharides (CO) are produced by arbuscular mycorrhizal fungi (AMF) and activate the plant symbiosis signalling pathway, which is essential for mycorrhiza formation. High-affinity LCO receptors belonging to the LysM receptor-like kinase (LysM-RLK) phylogenetic group LYR-IA play a role in AM establishment, but it is unclear which proteins are the plant high-affinity short-chain CO receptors. Here we studied members of the uncharacterized LYR-IB group, and found that they show high affinity for LCO, short- and long-chain CO, and play a complementary role with the LYR-IA receptors for AM establishment. While LYR-IB knock out mutants had a reduced AMF colonization in several species, constitutive/ectopic expression in wheat increased AMF colonization. LYR-IB function is conserved in all tested angiosperms, but in most japonica rice a deletion creates a frameshift in the gene, explaining differences in AM phenotypes between rice and other monocot single LYR-IA mutants. In conclusion, we identified a class of LysM-RLK receptors in angiosperms with unique biochemical properties and a role in both LCO and CO perception for AM establishment.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
- ANR-16-CE20-0025-01/Agence Nationale de la Recherche (French National Research Agency)
- ANR-10-LABX-41/Agence Nationale de la Recherche (French National Research Agency)
- ANR-18-EURE-0019/Agence Nationale de la Recherche (French National Research Agency)
- ANR-14-CE18-0008-01/Agence Nationale de la Recherche (French National Research Agency)
- ANR-14-CE18-0008-01/Agence Nationale de la Recherche (French National Research Agency)
- ANR-10-LABX-40-SPS/Agence Nationale de la Recherche (French National Research Agency)
- ANR-16-CE20-0025-01/Agence Nationale de la Recherche (French National Research Agency)
- ANR-17-EURE-0003/Agence Nationale de la Recherche (French National Research Agency)
- 32100241/National Natural Science Foundation of China (National Science Foundation of China)
LinkOut - more resources
Full Text Sources
