

ADCY4 inhibits cAMP-induced growth of breast cancer by inactivating FAK/AKT and ERK signaling but is frequently silenced by DNA methylation
- PMID: 40594405
- PMCID: PMC12217726
- DOI: 10.1038/s41598-025-06294-1
ADCY4 inhibits cAMP-induced growth of breast cancer by inactivating FAK/AKT and ERK signaling but is frequently silenced by DNA methylation
Abstract
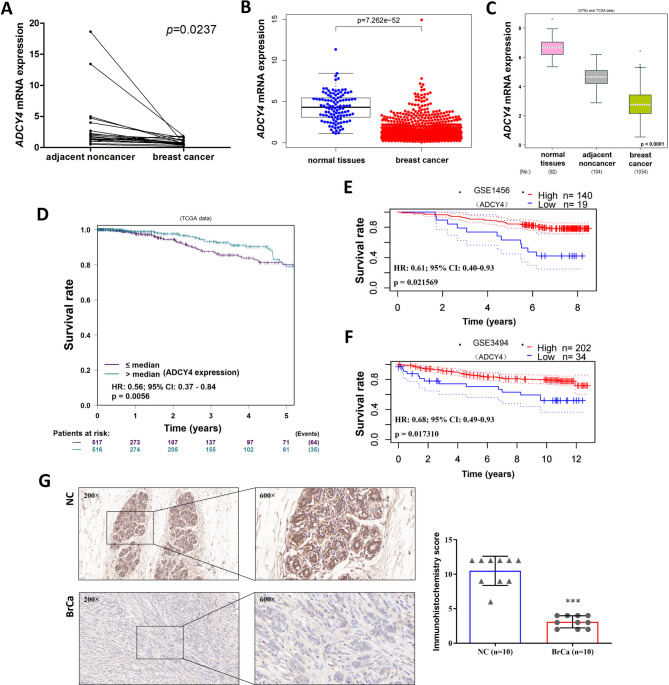
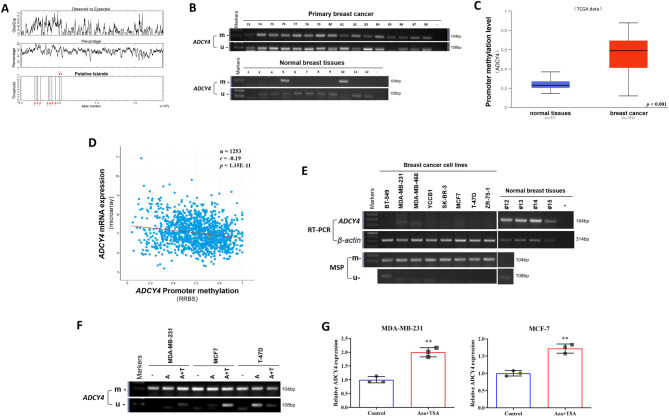
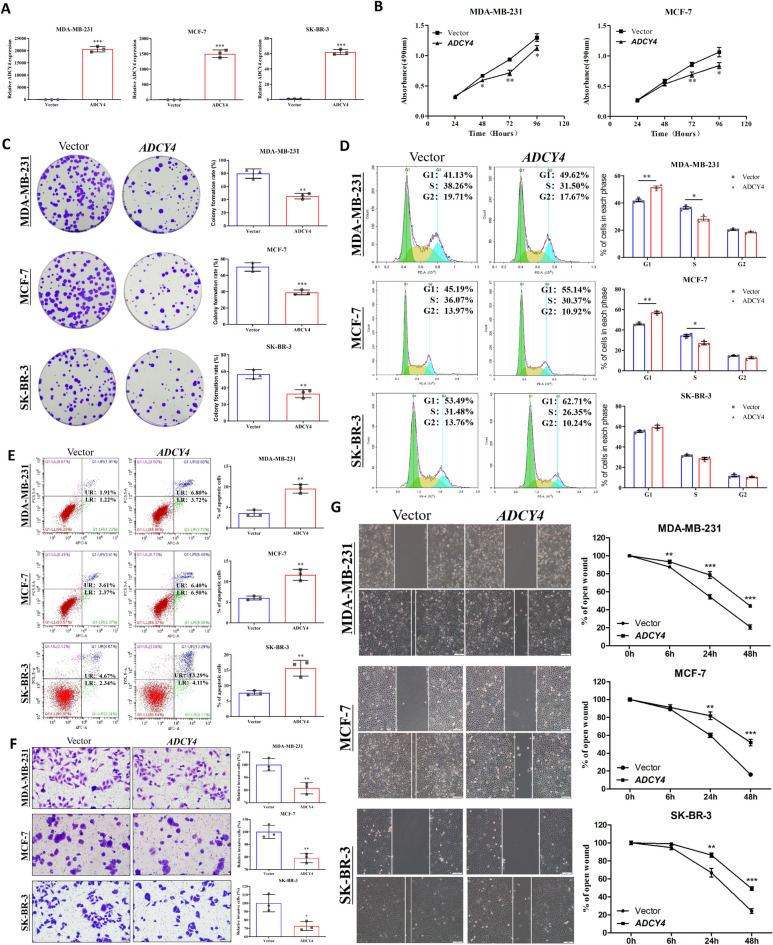
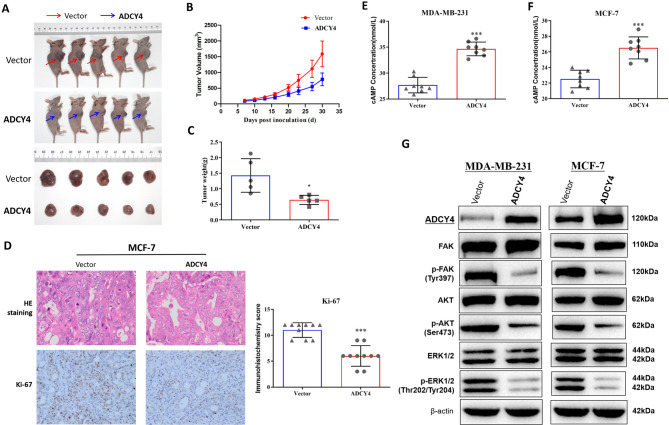
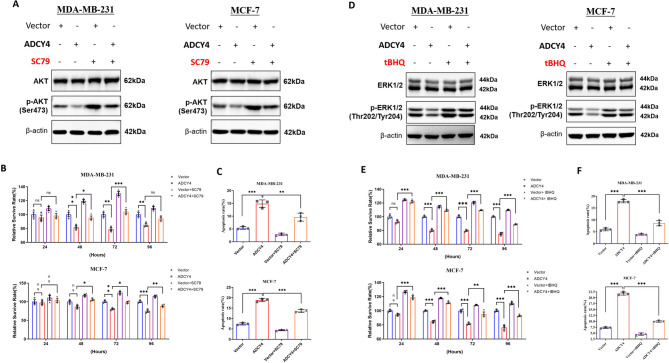
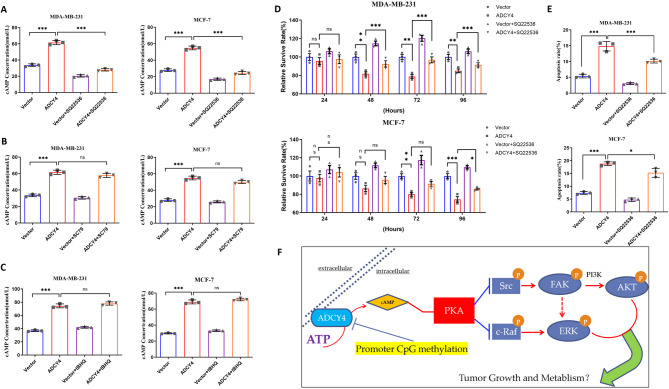
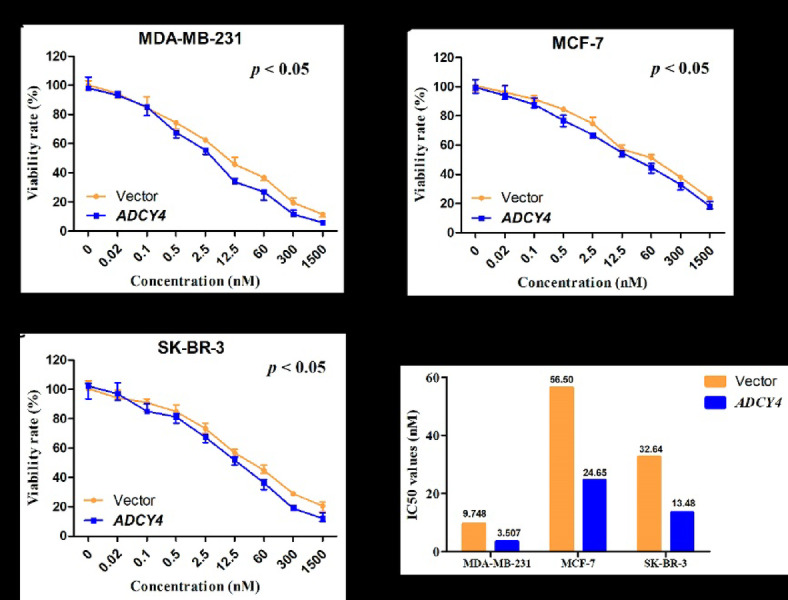
Local increases in cyclic adenosine monophosphate (cAMP) caused by specific adenylyl cyclases (ACs) can selectively modulate related proteins. AC-selective drugs have an advantage in side effect control, and the specific AC may finally be considered as a therapeutic target. We show that adenylyl cyclase 4 (ADCY4), which is silenced by DNA methylation and is critical for breast cancer (BC) patient survival, plays essential roles in anti-tumor effects in BC cells. DNA methyltransferase inhibitor and histone deacetylase inhibitor can restore ADCY4 mRNA expression in ADCY4-silenced BC cells. ADCY4 directly affects BC cell proliferation, apoptosis, invasion, and metastasis. Mechanistically, ADCY4 converts ATP to cAMP and activates cAMP/PKA signaling, leading to a decrease in the phosphorylation level of downstream FAK/AKT and ERK signaling and creating a suppression environment for cell survival. Ectopic ADCY4 inhibits BC growth, which is blocked by cAMP inhibition, activating AKT and ERK. The present study provides evidence that human BC relies upon this epigenetic silenced ADCY4-associated ATP-cAMP loop for phosphorylation and activation of FAK/AKT and ERK signaling. Also, ADCY4 increases BC cell chemosensitivity to paclitaxel. The observations demonstrates that ADCY4 is a significant tumor suppressor and that loss of ADCY4 functions by DNA methylation hampers cAMP signaling and triggers FAK/AKT and ERK signaling during breast tumorigenesis.
Keywords: ADCY4; Breast cancer; DNA methylation; FAK; cAMP.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests. Ethical approval and consent to participate: Ethics approval was received from the Affiliated Hospital of Chongqing Medical University. All participants provided written consent during their enrollment. Consent for publication: All patients provided written consent during enrollment for publication.
Figures
Similar articles
-
Upregulation of mTORC2 activation by the selective agonist of EPAC, 8-CPT-2Me-cAMP, in prostate cancer cells: assembly of a multiprotein signaling complex.J Cell Biochem. 2012 May;113(5):1488-500. doi: 10.1002/jcb.24018. J Cell Biochem. 2012. Retraction in: J Cell Biochem. 2025 Jul;126(7):e70048. doi: 10.1002/jcb.70048. PMID: 22173835 Retracted.
-
Upregulated SAE1 Drives Tumorigenesis and Is Associated with Poor Clinical Outcomes in Breast Cancer.Breast J. 2024 Jun 30;2024:2981722. doi: 10.1155/2024/2981722. eCollection 2024. Breast J. 2024. PMID: 39742381 Free PMC article.
-
Network-based meta-analysis and confirmation of genes ATP1A2, FXYD1, and ADCY3 associated with cAMP signaling in breast tumors compared to corresponding normal marginal tissues.Cell Mol Biol (Noisy-le-grand). 2024 Nov 27;70(11):16-30. doi: 10.14715/cmb/2024.70.11.3. Cell Mol Biol (Noisy-le-grand). 2024. PMID: 39707785
-
Blood-based DNA methylation as biomarker for breast cancer: a systematic review.Clin Epigenetics. 2016 Nov 14;8:115. doi: 10.1186/s13148-016-0282-6. eCollection 2016. Clin Epigenetics. 2016. PMID: 27895805 Free PMC article.
-
A rapid and systematic review of the clinical effectiveness and cost-effectiveness of paclitaxel, docetaxel, gemcitabine and vinorelbine in non-small-cell lung cancer.Health Technol Assess. 2001;5(32):1-195. doi: 10.3310/hta5320. Health Technol Assess. 2001. PMID: 12065068
References
-
- Sung, H. et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J. Clin.71, 209–249. 10.3322/caac.21660 (2021). - PubMed
-
- Loibl, S., Poortmans, P., Morrow, M., Denkert, C. & Curigliano, G. Breast cancer. Lancet397, 1750–1769. 10.1016/S0140-6736(20)32381-3 (2021). - PubMed
-
- Holubekova, V. et al. Epigenetic regulation by DNA methylation and MiRNA molecules in cancer. Future Oncol.13, 2217–2222. 10.2217/fon-2017-0363 (2017). - PubMed
-
- Pierre, S., Eschenhagen, T., Geisslinger, G. & Scholich, K. Capturing adenylyl cyclases as potential drug targets. Nat. Rev. Drug Discov.8, 321–335. 10.1038/nrd2827 (2009). - PubMed
-
- Dong, H., Claffey, K. P., Brocke, S. & Epstein, P. M. Inhibition of breast cancer cell migration by activation of cAMP signaling. Breast Cancer Res. Treat.152, 17–28. 10.1007/s10549-015-3445-9 (2015). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
