

Higher dopamine D1 receptor expression in prefrontal parvalbumin neurons underlies higher distractibility in marmosets versus macaques
- PMID: 40594842
- PMCID: PMC12214923
- DOI: 10.1038/s42003-025-08297-0
Higher dopamine D1 receptor expression in prefrontal parvalbumin neurons underlies higher distractibility in marmosets versus macaques
Abstract
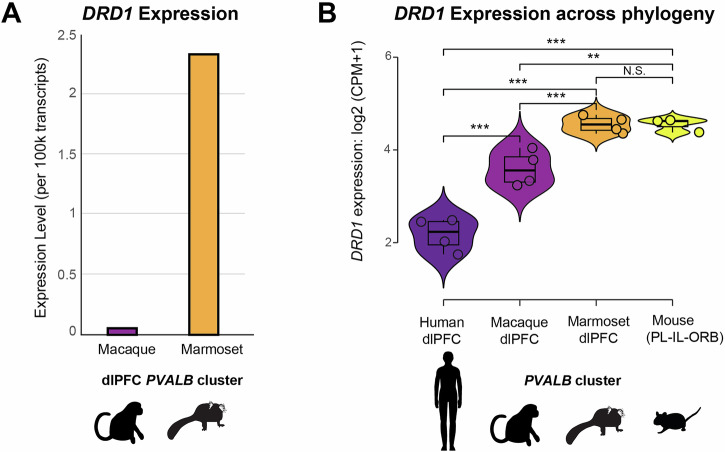
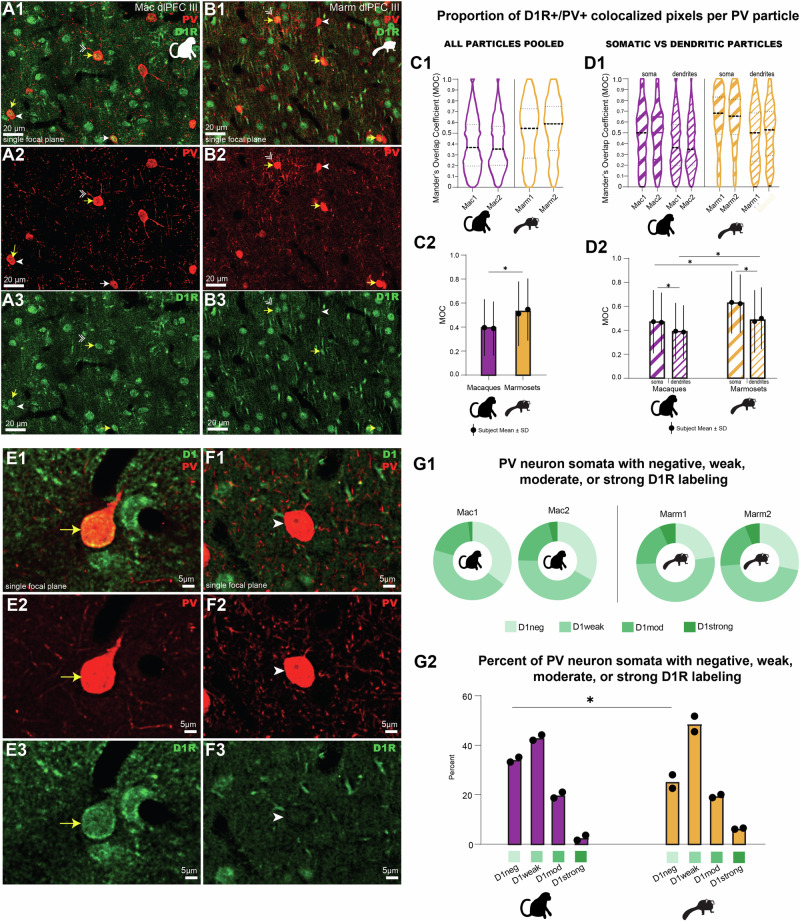
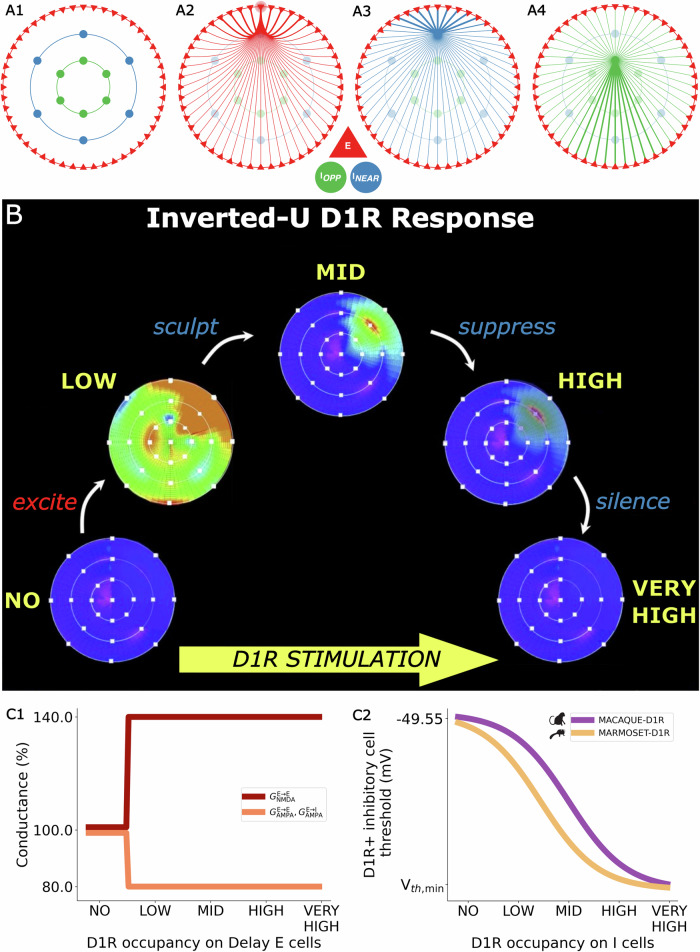
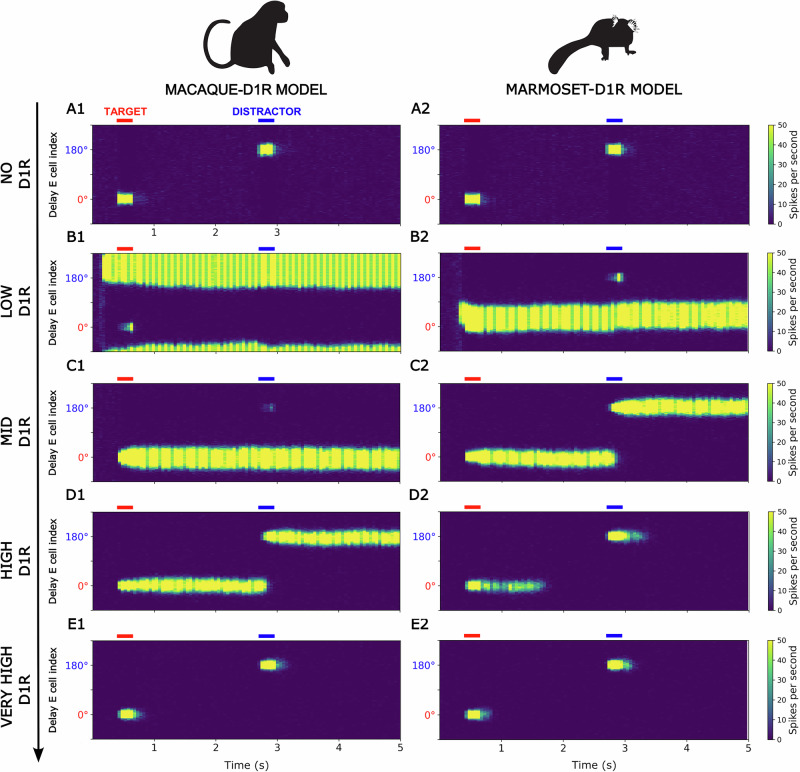
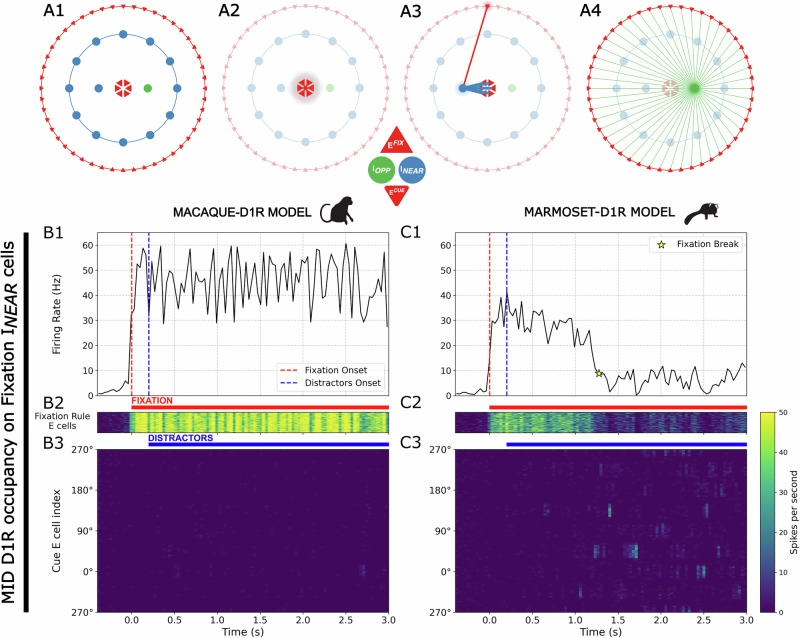
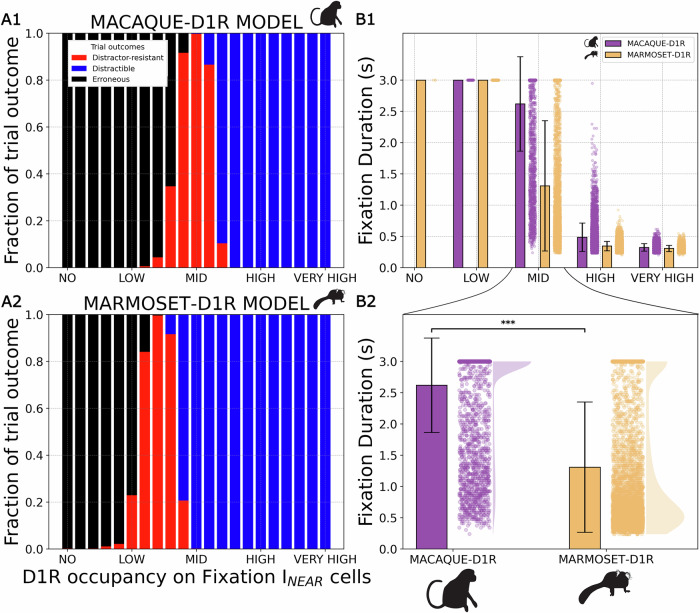
Marmosets and macaques are common nonhuman primate models of cognition, yet marmosets appear more distractible and perform worse in cognitive tasks. The dorsolateral prefrontal cortex (dlPFC) is pivotal for sustained attention, and research in macaques suggests that dopaminergic modulation and inhibitory parvalbumin (PV) neurons could influence distractor resistance. Here we compare the two species using a visual fixation task with distractors, perform molecular and anatomical analyses in dlPFC, and link functional microcircuitry with cognitive performance using computational modeling. We show that marmosets are more distractible than macaques, and that marmoset dlPFC PV neurons contain higher levels of dopamine D1 receptor (D1R) transcripts and protein, similar to levels in mice. Our modeling indicates that higher D1R expression in marmoset dlPFC PV neurons may increase distractibility by making dlPFC microcircuits more vulnerable to disruptions of their task-related persistent activity, especially when dopamine is released in dlPFC in response to unexpected salient stimuli.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
