

In situ structure of a bacterial flagellar motor at subnanometre resolution reveals adaptations for increased torque
- PMID: 40595286
- PMCID: PMC12222027
- DOI: 10.1038/s41564-025-02012-9
In situ structure of a bacterial flagellar motor at subnanometre resolution reveals adaptations for increased torque
Erratum in
-
Author Correction: In situ structure of a bacterial flagellar motor at subnanometre resolution reveals adaptations for increased torque.Nat Microbiol. 2025 Aug;10(8):2092. doi: 10.1038/s41564-025-02082-9. Nat Microbiol. 2025. PMID: 40659922 Free PMC article. No abstract available.
Abstract
The bacterial flagellar motor, which spins a helical propeller for propulsion, has undergone evolutionary diversification across bacterial species, often involving the addition of structures associated with increasing torque for motility in viscous environments. Understanding how such structures function and have evolved is hampered by challenges in visualizing motors in situ. Here we developed a Campylobacter jejuni minicell system for in situ cryogenic electron microscopy imaging and single-particle analysis of its motor, one of the most complex flagellar motors known, to subnanometre resolution. Focusing on the large periplasmic structures which are essential for increasing torque, our structural data, interpreted with molecular models, show that the basal disk comprises concentric rings of FlgP. The medial disk is a lattice of PflC with PflD, while the proximal disk is a rim of PflB attached to spokes of PflA. PflAB dimerization is essential for proximal disk assembly, recruiting FliL to scaffold more stator complexes at a wider radius which increases torque. We also acquired insights into universal principles of flagellar torque generation. This in situ approach is broadly applicable to other membrane-residing bacterial molecular machines.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
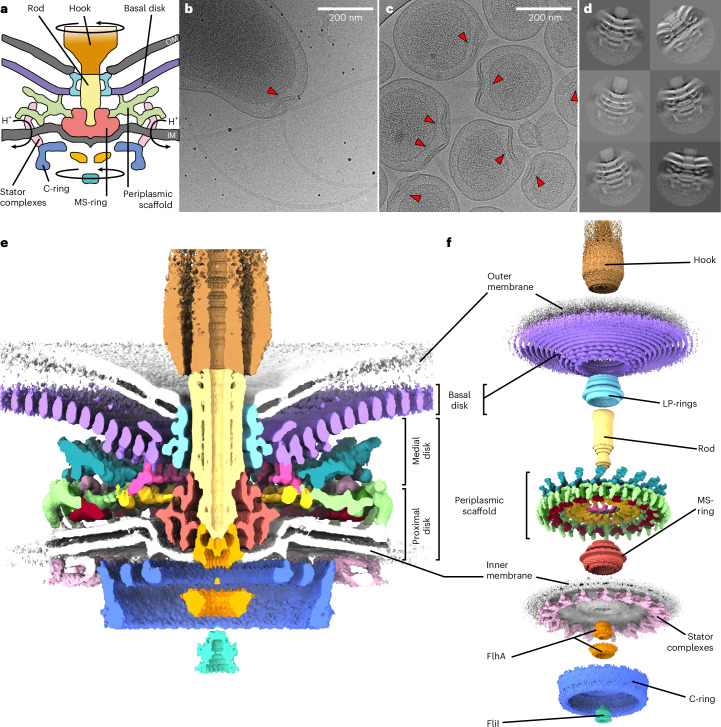
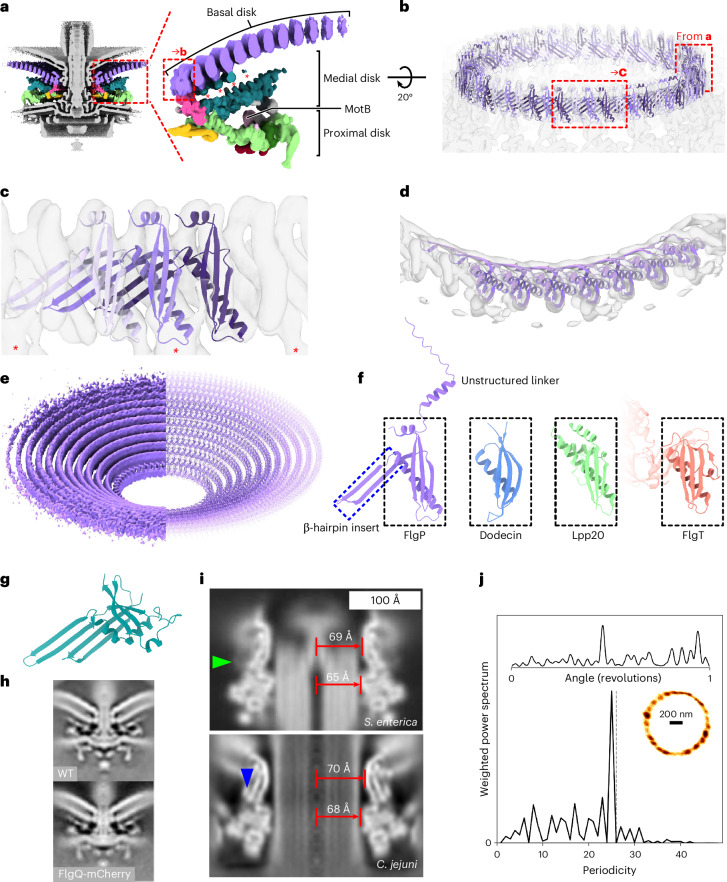
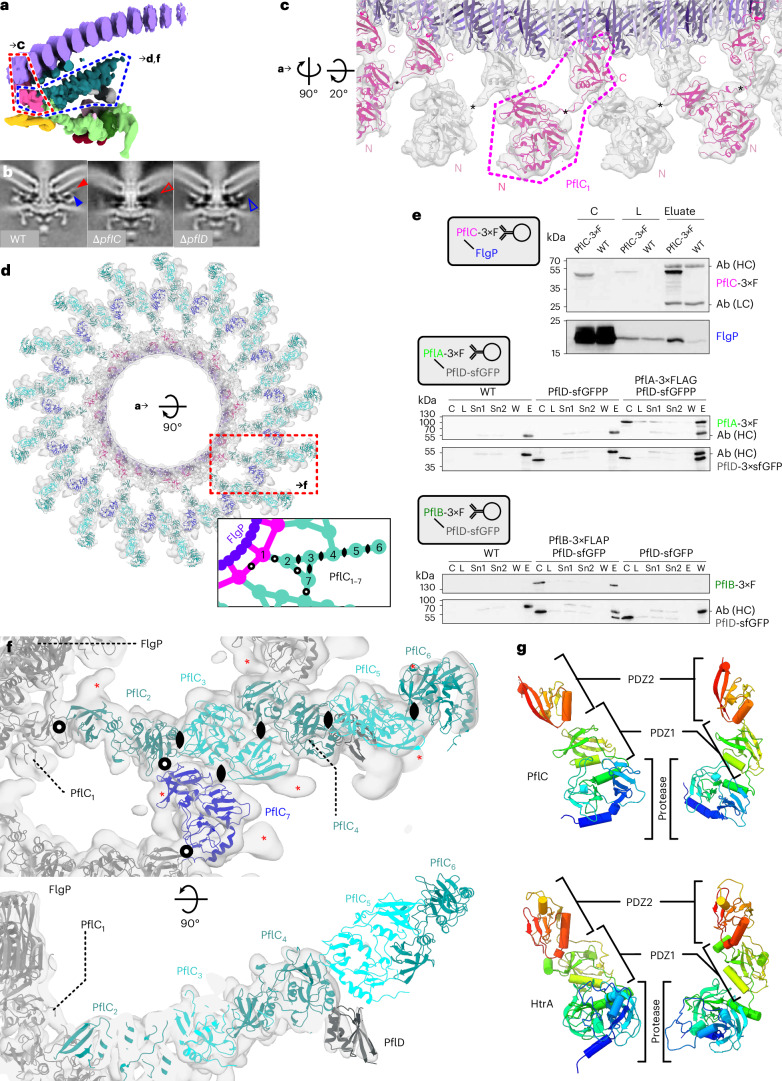
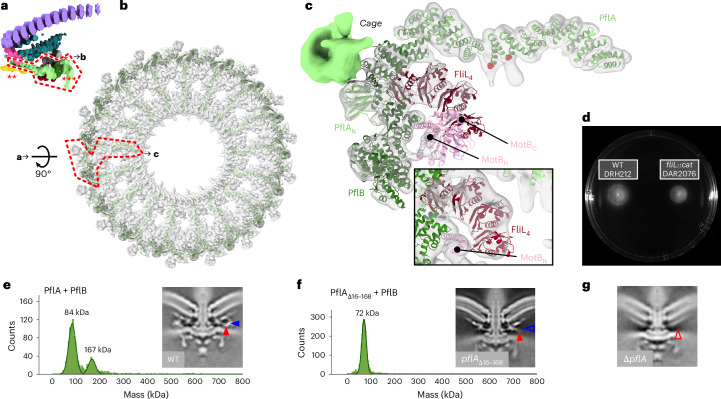
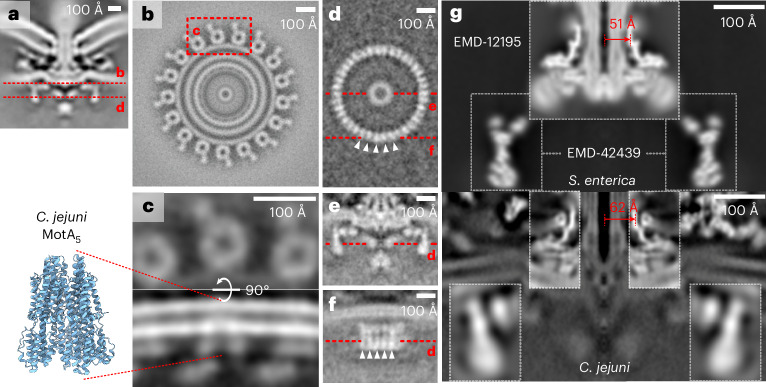
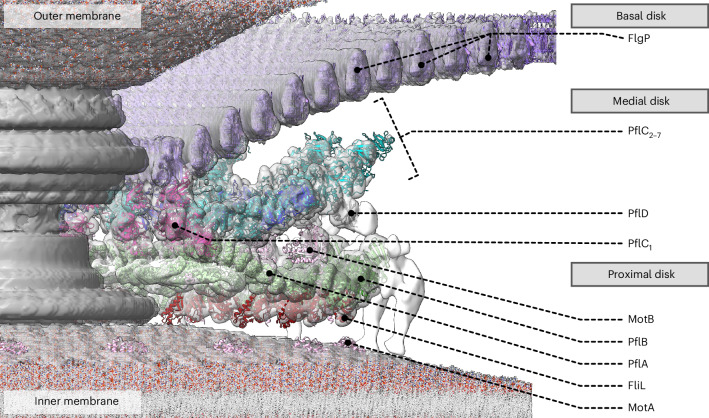


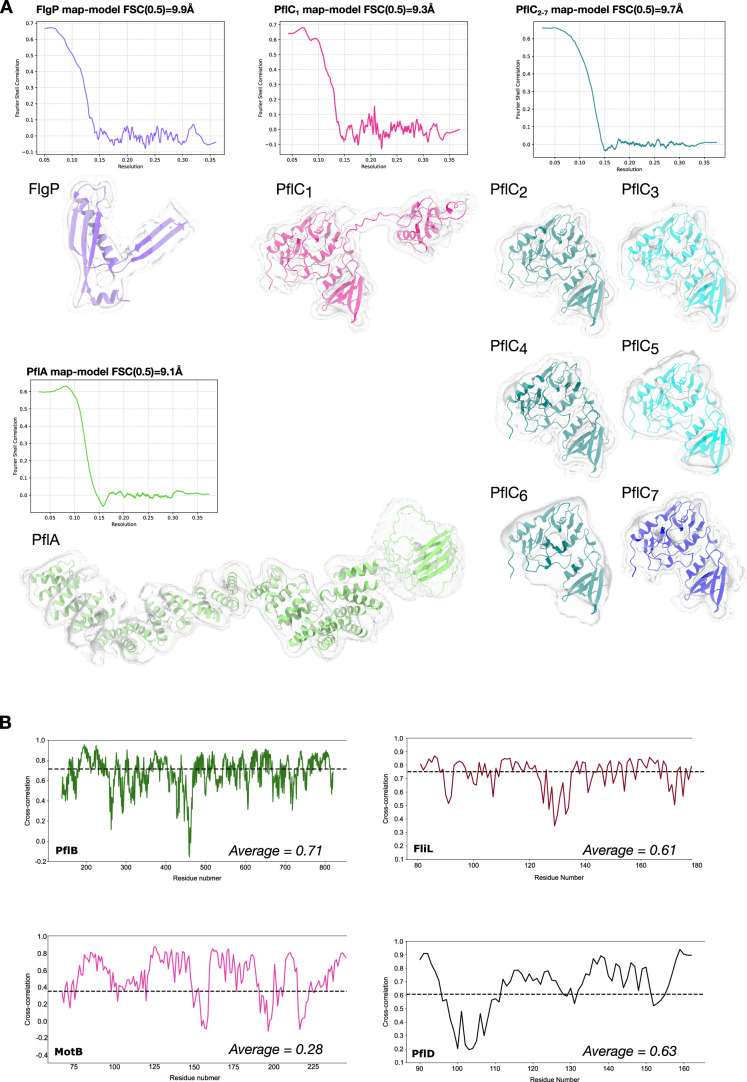






Update of
-
Molecular model of a bacterial flagellar motor in situ reveals a "parts-list" of protein adaptations to increase torque.bioRxiv [Preprint]. 2024 Oct 9:2023.09.08.556779. doi: 10.1101/2023.09.08.556779. bioRxiv. 2024. Update in: Nat Microbiol. 2025 Jul;10(7):1723-1740. doi: 10.1038/s41564-025-02012-9. PMID: 39416179 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- Deutscher Akademischer Austauschdienst (German Academic Exchange Service)
- BB/M011178/1/RCUK | Biotechnology and Biological Sciences Research Council (BBSRC)
- ANR-10-INBS-04-01/Agence Nationale de la Recherche (French National Research Agency)
- RGP0028/2021-HOCHBERG/Human Frontier Science Program (HFSP)
- CC2106/RCUK | Medical Research Council (MRC)
- ANR-10-INBS-05/Agence Nationale de la Recherche (French National Research Agency)
- Sh580/7-2/Deutsche Forschungsgemeinschaft (German Research Foundation)
- NIH R01 AI065539/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- PHYBION ANR-23-ERCB-0005-01/Agence Nationale de la Recherche (French National Research Agency)
- CC2106/Wellcome Trust (Wellcome)
- MR/V000799/1/RCUK | Medical Research Council (MRC)
- WT_/Wellcome Trust/United Kingdom
- CC2106/Cancer Research UK (CRUK)
- R01 AI065539/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
