

Stable recurrent dynamics in heterogeneous neuromorphic computing systems using excitatory and inhibitory plasticity
- PMID: 40595529
- PMCID: PMC12214931
- DOI: 10.1038/s41467-025-60697-2
Stable recurrent dynamics in heterogeneous neuromorphic computing systems using excitatory and inhibitory plasticity
Abstract
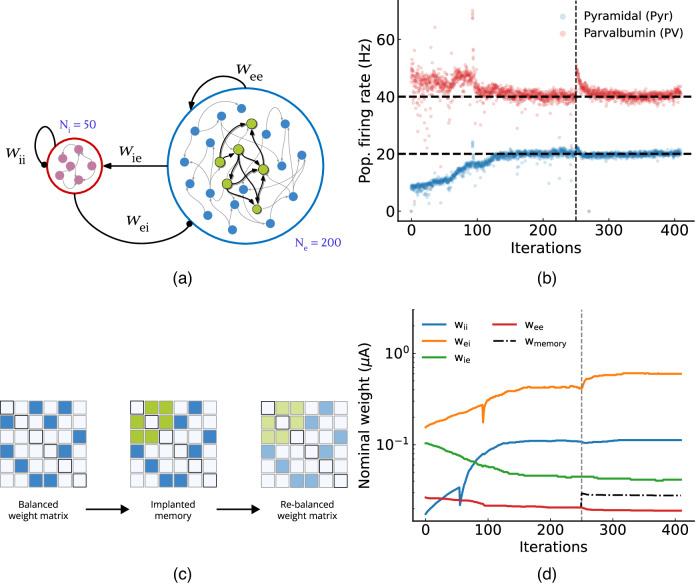
Many neural computations emerge from self-sustained patterns of activity in recurrent neural circuits, which rely on balanced excitation and inhibition. Neuromorphic electronic circuits represent a promising approach for implementing the brain's computational primitives. However, achieving the same robustness of biological networks in neuromorphic systems remains a challenge due to the variability in their analog components. Inspired by real cortical networks, we apply a biologically-plausible cross-homeostatic rule to balance neuromorphic implementations of spiking recurrent networks. We demonstrate how this rule can autonomously tune the network to produce robust, self-sustained dynamics in an inhibition-stabilized regime, even in presence of device mismatch. It can implement multiple, co-existing stable memories, with emergent soft-winner-take-all and reproduce the "paradoxical effect" observed in cortical circuits. In addition to validating neuroscience models on a substrate sharing many similar limitations with biological systems, this enables the automatic configuration of ultra-low power, mixed-signal neuromorphic technologies despite the large chip-to-chip variability.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures







Similar articles
-
Paradoxical self-sustained dynamics emerge from orchestrated excitatory and inhibitory homeostatic plasticity rules.Proc Natl Acad Sci U S A. 2022 Oct 25;119(43):e2200621119. doi: 10.1073/pnas.2200621119. Epub 2022 Oct 17. Proc Natl Acad Sci U S A. 2022. PMID: 36251988 Free PMC article.
-
Selective inhibition in CA3: A mechanism for stable pattern completion through heterosynaptic plasticity.PLoS Comput Biol. 2025 Jul 7;21(7):e1013267. doi: 10.1371/journal.pcbi.1013267. eCollection 2025 Jul. PLoS Comput Biol. 2025. PMID: 40623085 Free PMC article.
-
Biologically-informed excitatory and inhibitory ratio for robust spiking neural network training.Sci Rep. 2025 Jul 10;15(1):24798. doi: 10.1038/s41598-025-03408-7. Sci Rep. 2025. PMID: 40634353 Free PMC article.
-
Spiking Neural Networks for Multimodal Neuroimaging: A Comprehensive Review of Current Trends and the NeuCube Brain-Inspired Architecture.Bioengineering (Basel). 2025 Jun 9;12(6):628. doi: 10.3390/bioengineering12060628. Bioengineering (Basel). 2025. PMID: 40564444 Free PMC article. Review.
-
Dissociated neuronal cultures as model systems for self-organized prediction.Front Neural Circuits. 2025 Jun 25;19:1568652. doi: 10.3389/fncir.2025.1568652. eCollection 2025. Front Neural Circuits. 2025. PMID: 40635884 Free PMC article. Review.
References
-
- Buonomano, D. V. & Maass, W. State-dependent computations: spatiotemporal processing in cortical networks. Nat. Rev. Neurosci.10, 113–125 (2009). - PubMed
-
- Douglas, R. J. & Martin, K. A. Recurrent neuronal circuits in the neocortex. Curr. Biol.17, R496–R500 (2007). - PubMed
-
- Douglas, R. J., Koch, C., Mahowald, M., Martin, K. A. C. & Suarez, H. H. Recurrent excitation in neocortical circuits. Science269, 981–985 (1995). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
