

Genomic landscape of virus-associated cancers
- PMID: 40595559
- PMCID: PMC12219571
- DOI: 10.1038/s41467-025-60836-9
Genomic landscape of virus-associated cancers
Abstract
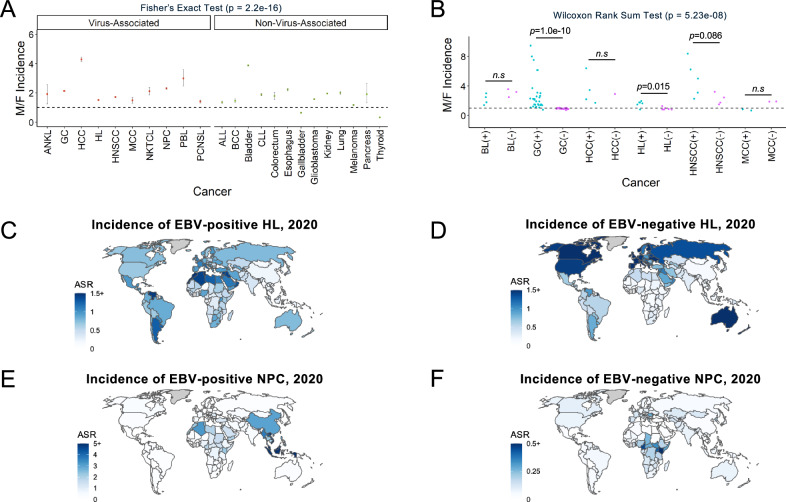
It has been estimated that 15%-20% of human cancers are attributable to infections, mostly by carcinogenic viruses. The incidence varies worldwide, with a majority affecting developing countries. Here, we conduct a comparative analysis of virus-positive and virus-negative tumors in nine cancers linked to five viruses. We observe a higher frequency of virus-positive tumors in males, with notable geographic differences in incidence. Our genomic analysis of 1971 tumors reveals a lower somatic burden, distinct mutation signatures, and driver gene mutations in virus-positive tumors. Compared to virus-negative cases, virus-positive cases have fewer mutations of TP53, CDKN2A, and deletions of 9p21.3/CDKN2A-CDKN1A while exhibiting more mutations in RNA helicases DDX3X and EIF4A1. Furthermore, an analysis of clinical trials of PD-(L)1 inhibitors suggests an association of virus-positivity with higher treatment response rate, particularly evident in gastric cancer and head and neck squamous cell carcinoma. Both cancer types also show evidence of increased CD8 + T cell infiltration and T cell receptor clonal selection in virus-positive tumors. These results illustrate the epidemiological, genetic, and therapeutic trends across virus-associated malignancies.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: R. Rabadan is the founder of Genotwin, a member of the advisory board of Diatech Pharmacogenetics and Flahy. None of these activities are related to the results in the current manuscript. The remaining authors declare no competing interests.
Figures





Update of
-
Genomic landscape of virus-associated cancers.medRxiv [Preprint]. 2023 Feb 16:2023.02.14.23285775. doi: 10.1101/2023.02.14.23285775. medRxiv. 2023. Update in: Nat Commun. 2025 Jul 1;16(1):5887. doi: 10.1038/s41467-025-60836-9. PMID: 36824731 Free PMC article. Updated. Preprint.
References
-
- de Martel, C., Georges, D., Bray, F., Ferlay, J. & Clifford, G. M. Global burden of cancer attributable to infections in 2018: a worldwide incidence analysis. Lancet Glob. Health8, e180–e190 (2020). - PubMed
-
- Schrama, D. et al. Merkel cell polyomavirus status is not associated with clinical course of Merkel cell carcinoma. J. Invest. Dermatol.131, 1631–1638 (2011). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
