

Lycorine hydrochloride inhibits cholangiocarcinoma through cholesterol biosynthesis and PTPN11 nuclear translocation
- PMID: 40598214
- PMCID: PMC12220003
- DOI: 10.1186/s12964-025-02318-5
Lycorine hydrochloride inhibits cholangiocarcinoma through cholesterol biosynthesis and PTPN11 nuclear translocation
Abstract
Background: Intrahepatic Cholangiocarcinoma (ICC) is a highly aggressive malignancy with limited treatment options. Identifying novel therapeutic agents for ICC is crucial. Numerous natural compounds have demonstrated remarkable anti-tumor activities and can enhance the efficacy of chemotherapy. Thus, our study aimed to screen natural compounds for their anti-ICC effects.
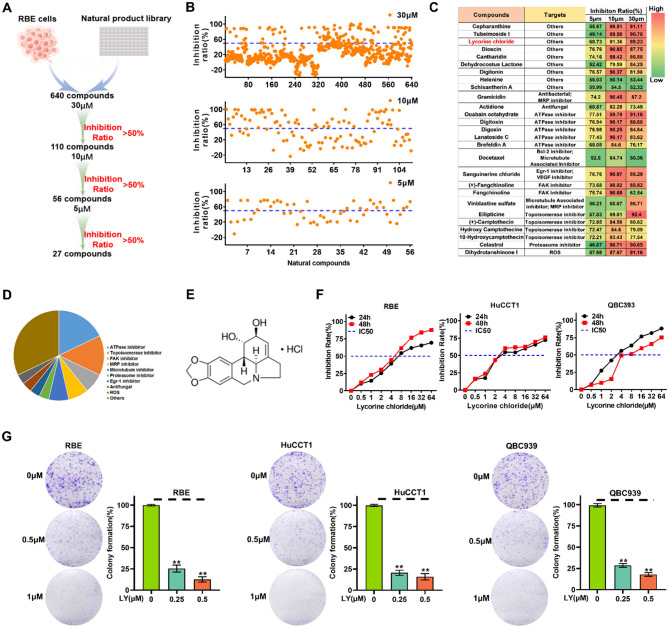
Methods: A total of 640 natural compounds were screened using a cell viability assay to identify potential compounds that could inhibit the proliferation of ICC cells. The anti-ICC effects of Lycorine Hydrochloride (LY) were confirmed through cell proliferation, colony formation, cell cycle, migration, and invasion assays, as well as in xenograft models. Bioinformatics analyses and validation experiments (Quantitative real-time PCR, Western blot, and immunostaining assays) were utilized to investigate the roles of genes (SQLE, FDFT1, and PTPN11) in ICC. RNA sequencing and immunofluorescence staining were performed to elucidate underlying molecular mechanisms.
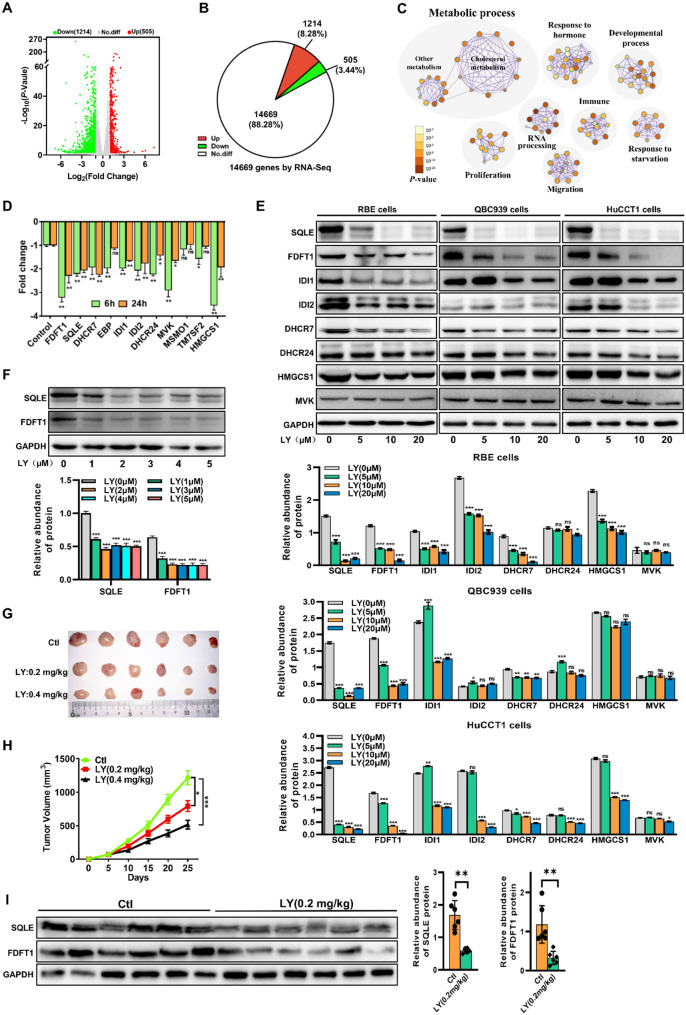
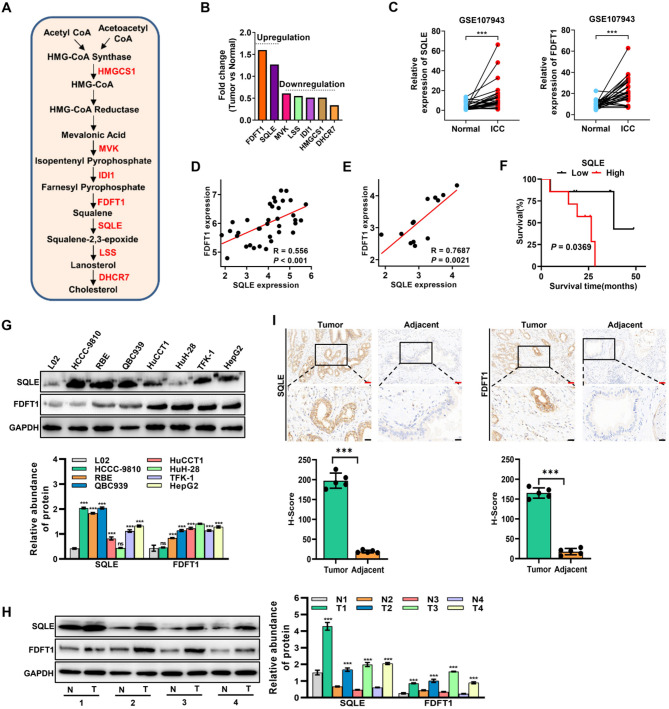
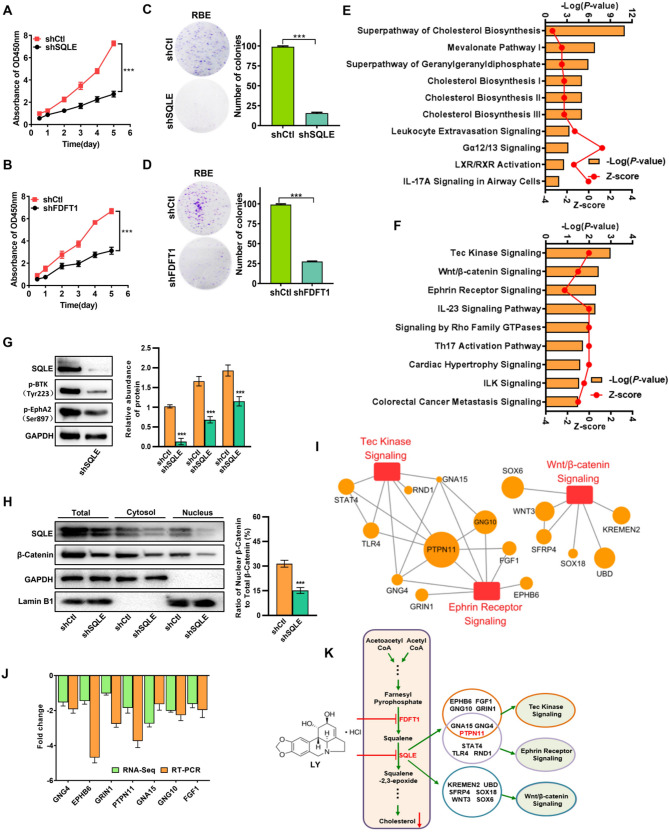
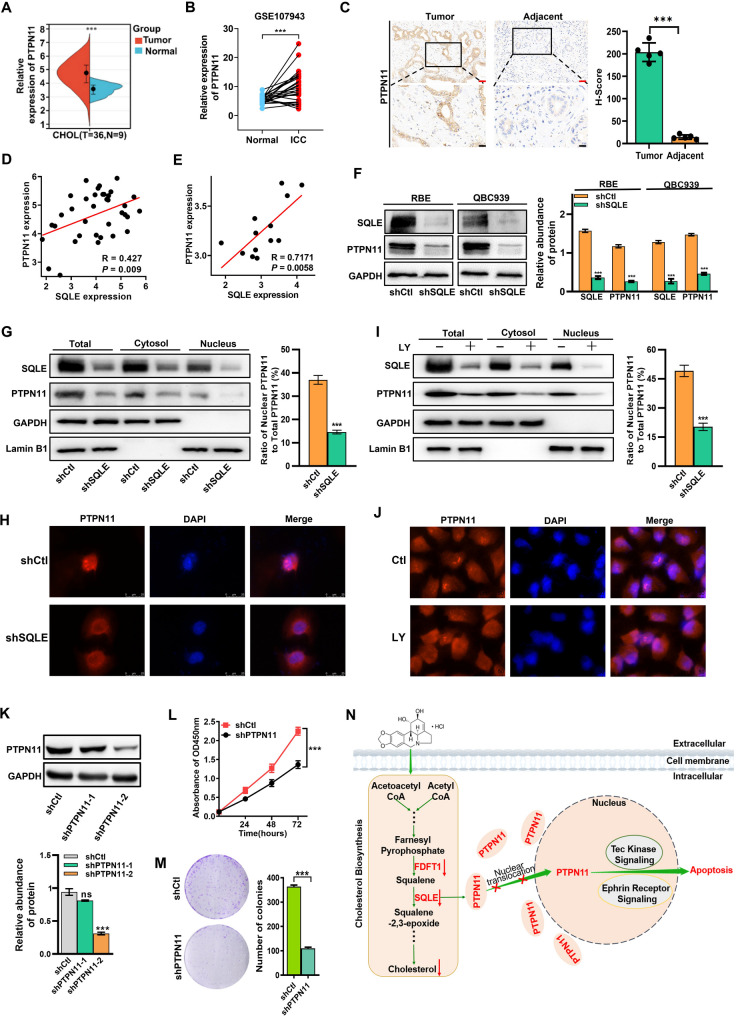
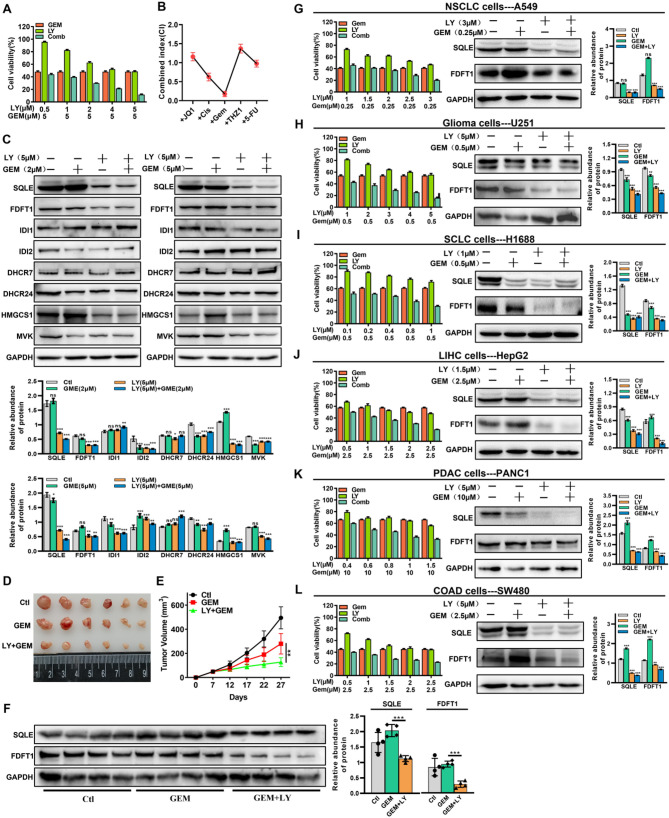
Results: LY was identified as a potential ICC inhibitor, exhibiting anti-ICC effects both in vitro and in vivo. Mechanistically, LY inhibited cholesterol synthesis in tumor cells by down-regulating the expression of SQLE and FDFT1. The knockdown of SQLE or FDFT1 significantly inhibited ICC cell proliferation and colony formation. RNA sequencing confirmed that inhibition of FDFT1 suppressed the cholesterol biosynthesis pathway, while SQLE inhibition affected specific oncogenic pathways. Additionally, immunofluorescence staining revealed that down-regulation of SQLE reduced PTPN11 expression and inhibited its nuclear translocation. Furthermore, pharmacological inhibition of SQLE and FDFT1 by LY significantly enhanced sensitivity to several common chemotherapeutic drugs for ICC. Notably, the combination of LY and Gemcitabine (GEM) displayed the most potent synergistic anti-tumor effect across various tumor types.
Conclusion: These findings identify Lycorine Hydrochloride as a promising treatment alternative for ICC and propose a novel combination strategy (LY + GEM) for treating multiple solid tumors.
Keywords: Cholangiocarcinoma; Cholesterol biosynthesis; FDFT1; Lycorine Hydrochloride; SQLE.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethical approval: The protocol and any procedures involving the care and use of animals in this study were reviewed and approved by the Institutional Animal Care and Use Committee of Zhongshan City People’s Hospital. All the paraffin-embedded tissues of patients used in this study were obtained by informed patient consent. Competing interests: The authors declare no competing interests.
Figures
Similar articles
-
Circ_0084927 promotes progression of intrahepatic cholangiocarcinoma by sponging miR-4725-5p to activate the PDPK1/AKT/mTOR signaling pathway.Cell Signal. 2025 Oct;134:111965. doi: 10.1016/j.cellsig.2025.111965. Epub 2025 Jun 26. Cell Signal. 2025. PMID: 40581264
-
PTP4A1 promotes intrahepatic cholangiocarcinoma development and progression by interacting with PTEN and activating the PI3K/AKT/GSKα axis.Oncol Rep. 2025 Oct;54(4):125. doi: 10.3892/or.2025.8958. Epub 2025 Aug 1. Oncol Rep. 2025. PMID: 40747713 Free PMC article.
-
The antitumor effects of lupenone on colon cancer and its mechanistic insights.Phytomedicine. 2025 Sep;145:156939. doi: 10.1016/j.phymed.2025.156939. Epub 2025 Jun 2. Phytomedicine. 2025. PMID: 40669211
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
References
-
- Moris D, Palta M, Kim C, Allen PJ, Morse MA, Lidsky ME. Advances in the treatment of intrahepatic cholangiocarcinoma: an overview of the current and future therapeutic landscape for clinicians. CA Cancer J Clin. 2023;73:198–222. - PubMed
-
- Valle J, Wasan H, Palmer DH, Cunningham D, Anthoney A, Maraveyas A, et al. Cisplatin plus gemcitabine versus gemcitabine for biliary tract cancer. N Engl J Med. 2010;362:1273–81. - PubMed
-
- Kelley RK, Bridgewater J, Gores GJ, Zhu AX. Systemic therapies for intrahepatic cholangiocarcinoma. J Hepatol. 2020;72:353–63. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
