

Harnessing the analog computing power of regulatory networks with the Regulatory Network Machine
- PMID: 40600146
- PMCID: PMC12210318
- DOI: 10.1016/j.isci.2025.112536
Harnessing the analog computing power of regulatory networks with the Regulatory Network Machine
Abstract
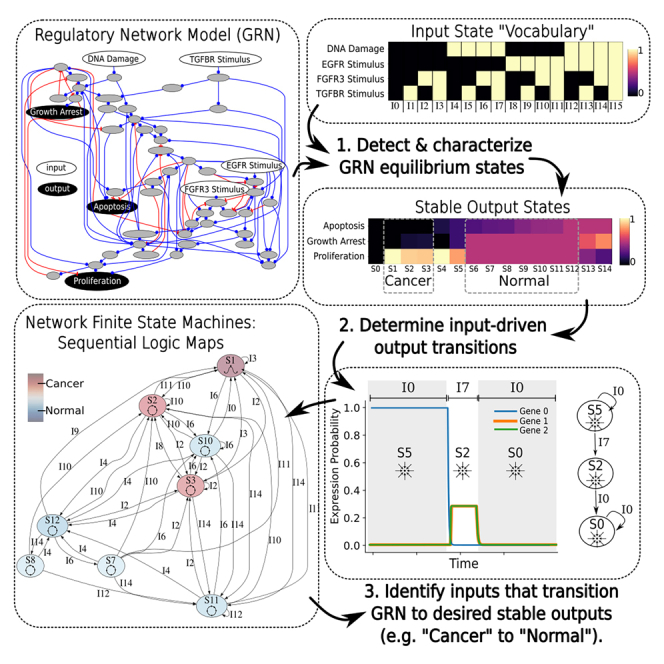
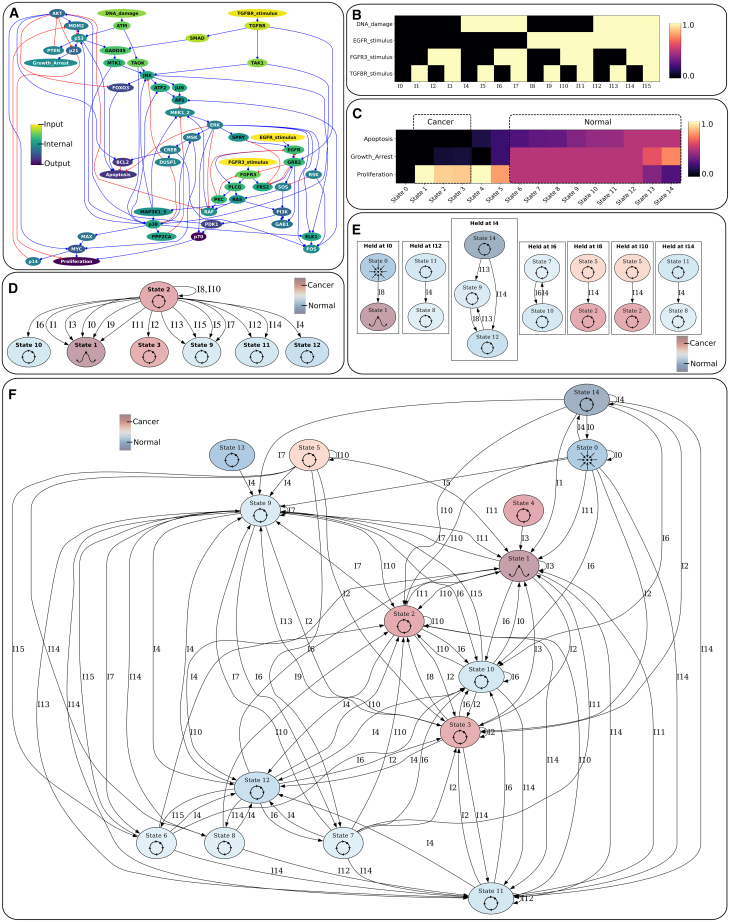
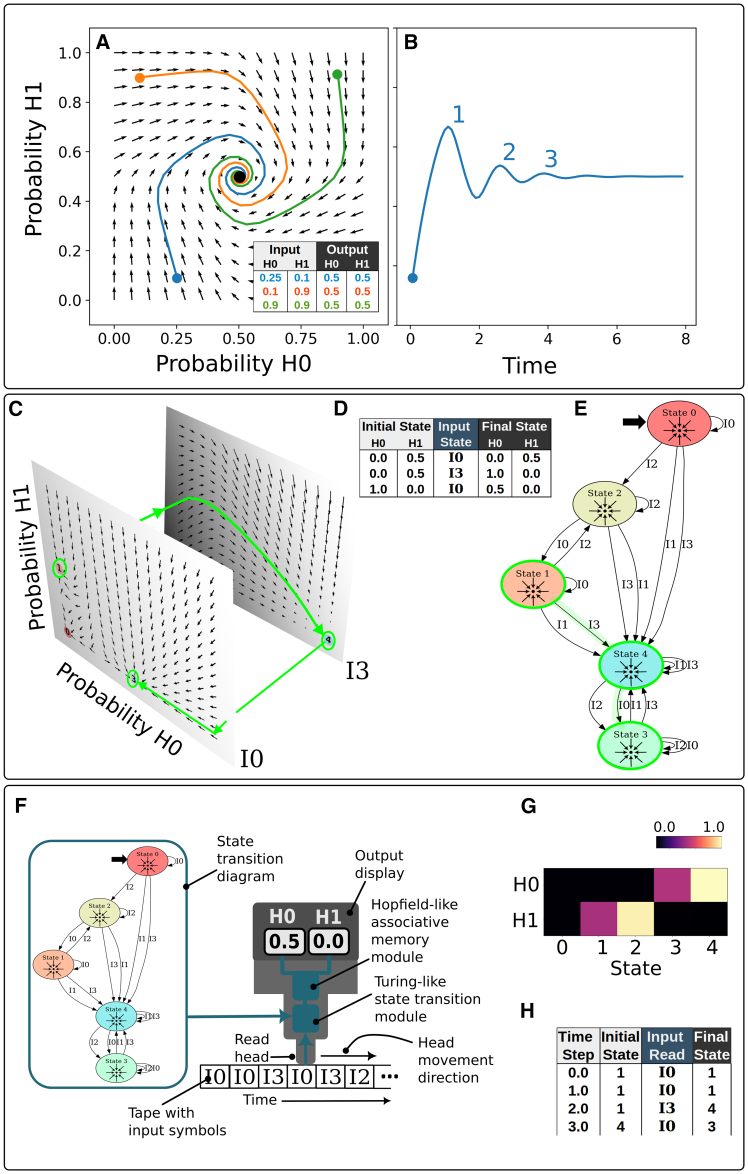
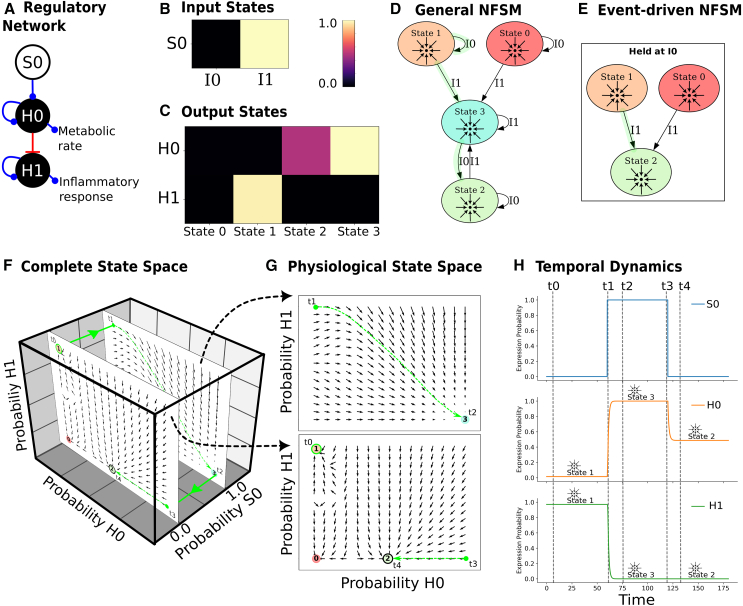
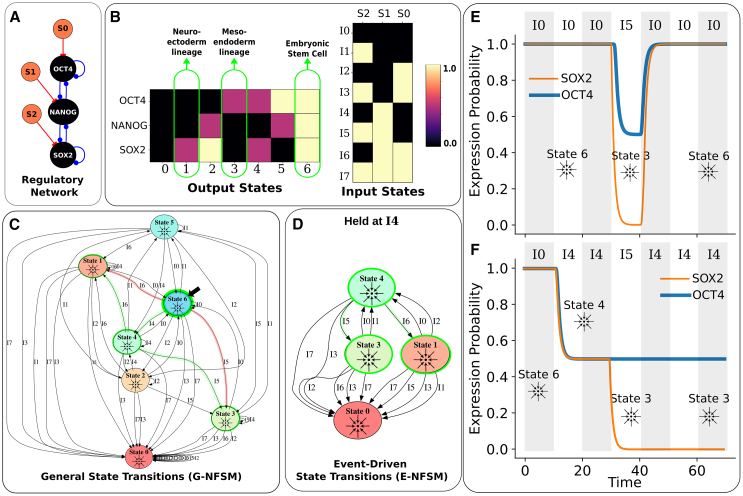
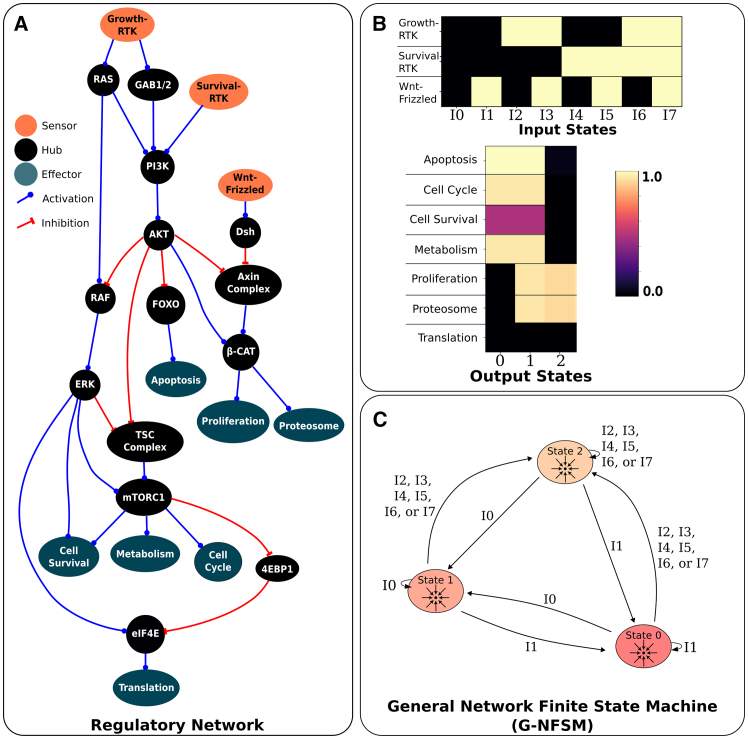
Gene regulatory networks (GRNs) are critically important for efforts in biomedicine and biotechnology. Here, we introduce the Regulatory Network Machine (RNM) framework, demonstrating how GRNs behave as analog computers capable of sophisticated information processing. Our RNM framework encapsulates: (1) a dissipative dynamic system with a focus on GRNs, (2) a set of inputs to the system, (3) system output states with identifiable relevance to biotechnological or biomedical objectives, and (4) Network Finite State Machines (NFSMs), which are maps detailing how the system changes equilibrium state in response to patterns of applied inputs. As an extension to attractor landscape analysis, the NFSMs map the sequential logic inherent in the GRN and, therefore, embody the "software-like" nature of the system, providing easy identification of specific applied interventions necessary to achieve desired, stable biological outcomes. We illustrate the use of our RNM framework in important biological examples, including in cancer renormalization.
Keywords: Bioengineering; Computer science; Systems biology.
© 2025 The Authors.
Conflict of interest statement
M.L. is scientific co-founder and minor shareholder of Astonishing Labs, a company seeking to develop advances in regenerative medicine based on proto-cognitive properties of gene-regulatory networks. Astonishing Labs provides a sponsored research agreement to Tufts University to support this research. M.L. also has an associate faculty appointment at Harvard’s Wyss Institute. M.L. and A.P. are listed as co-inventors on a provisional patent application covering the work reported here.
Figures





Similar articles
-
Factors that impact on the use of mechanical ventilation weaning protocols in critically ill adults and children: a qualitative evidence-synthesis.Cochrane Database Syst Rev. 2016 Oct 4;10(10):CD011812. doi: 10.1002/14651858.CD011812.pub2. Cochrane Database Syst Rev. 2016. PMID: 27699783 Free PMC article.
-
The health economics of insulin therapy: How do we address the rising demands, costs, inequalities and barriers to achieving optimal outcomes.Diabetes Obes Metab. 2025 Jul;27 Suppl 5(Suppl 5):24-35. doi: 10.1111/dom.16488. Epub 2025 Jun 4. Diabetes Obes Metab. 2025. PMID: 40464081 Free PMC article.
-
Surgical interventions for treating extracapsular hip fractures in older adults: a network meta-analysis.Cochrane Database Syst Rev. 2022 Feb 10;2(2):CD013405. doi: 10.1002/14651858.CD013405.pub2. Cochrane Database Syst Rev. 2022. PMID: 35142366 Free PMC article.
-
How to Implement Digital Clinical Consultations in UK Maternity Care: the ARM@DA Realist Review.Health Soc Care Deliv Res. 2025 May;13(22):1-77. doi: 10.3310/WQFV7425. Health Soc Care Deliv Res. 2025. PMID: 40417997 Review.
-
Direct-acting antivirals for chronic hepatitis C.Cochrane Database Syst Rev. 2017 Sep 18;9(9):CD012143. doi: 10.1002/14651858.CD012143.pub3. Cochrane Database Syst Rev. 2017. PMID: 28922704 Free PMC article.
References
-
- Lyon P. The biogenic approach to cognition. Cogn. Process. 2006;7:11–29. - PubMed
-
- Katz Y., Springer M., Fontana W. Embodying probabilistic inference in biochemical circuits [Internet] arXiv. 2018 http://arxiv.org/abs/1806.10161 Preprint at.
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
