

Multi-omic analysis reveals transkingdom gut dysbiosis in metabolic dysfunction-associated steatotic liver disease
- PMID: 40604156
- PMCID: PMC12320953
- DOI: 10.1038/s42255-025-01318-6
Multi-omic analysis reveals transkingdom gut dysbiosis in metabolic dysfunction-associated steatotic liver disease
Abstract
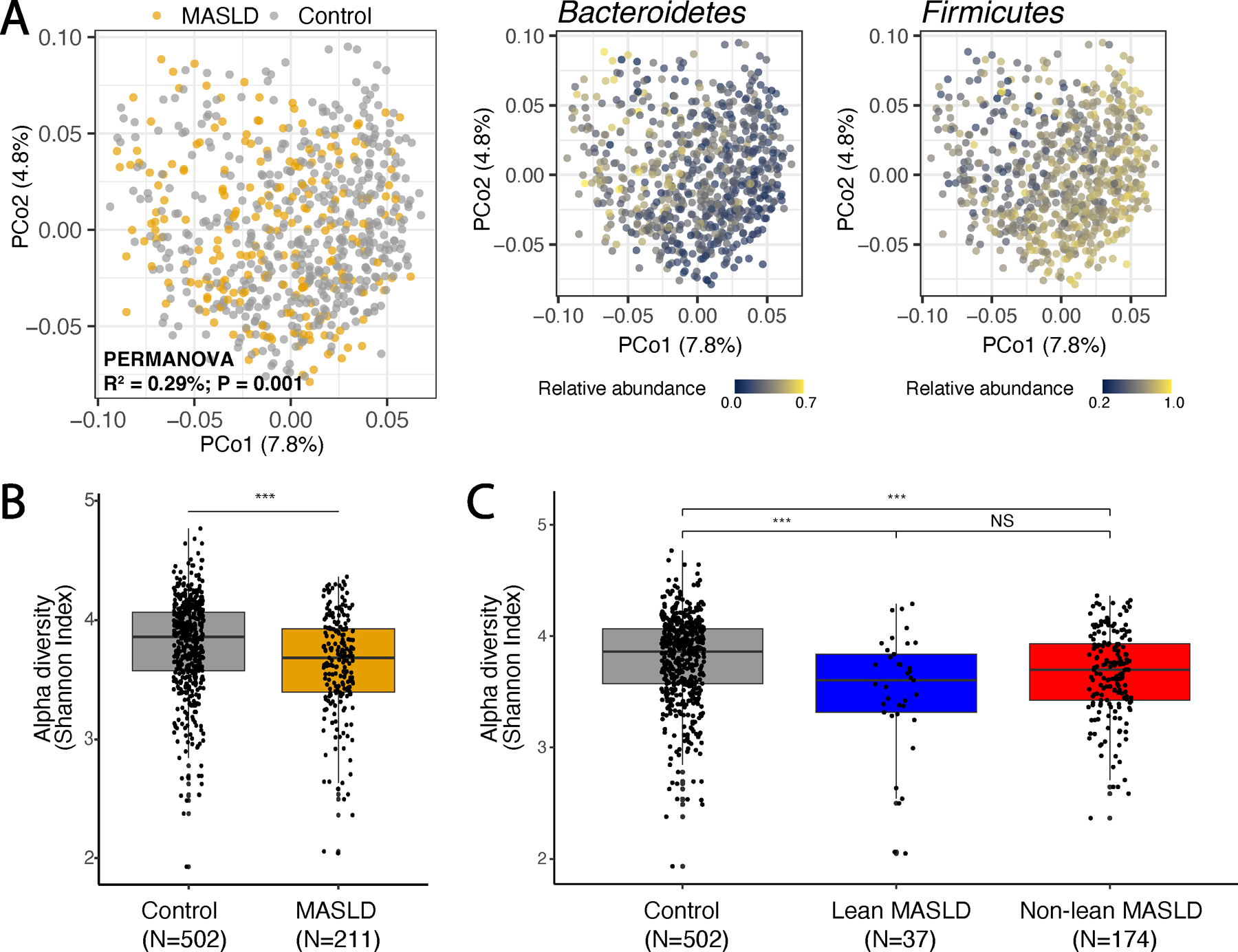
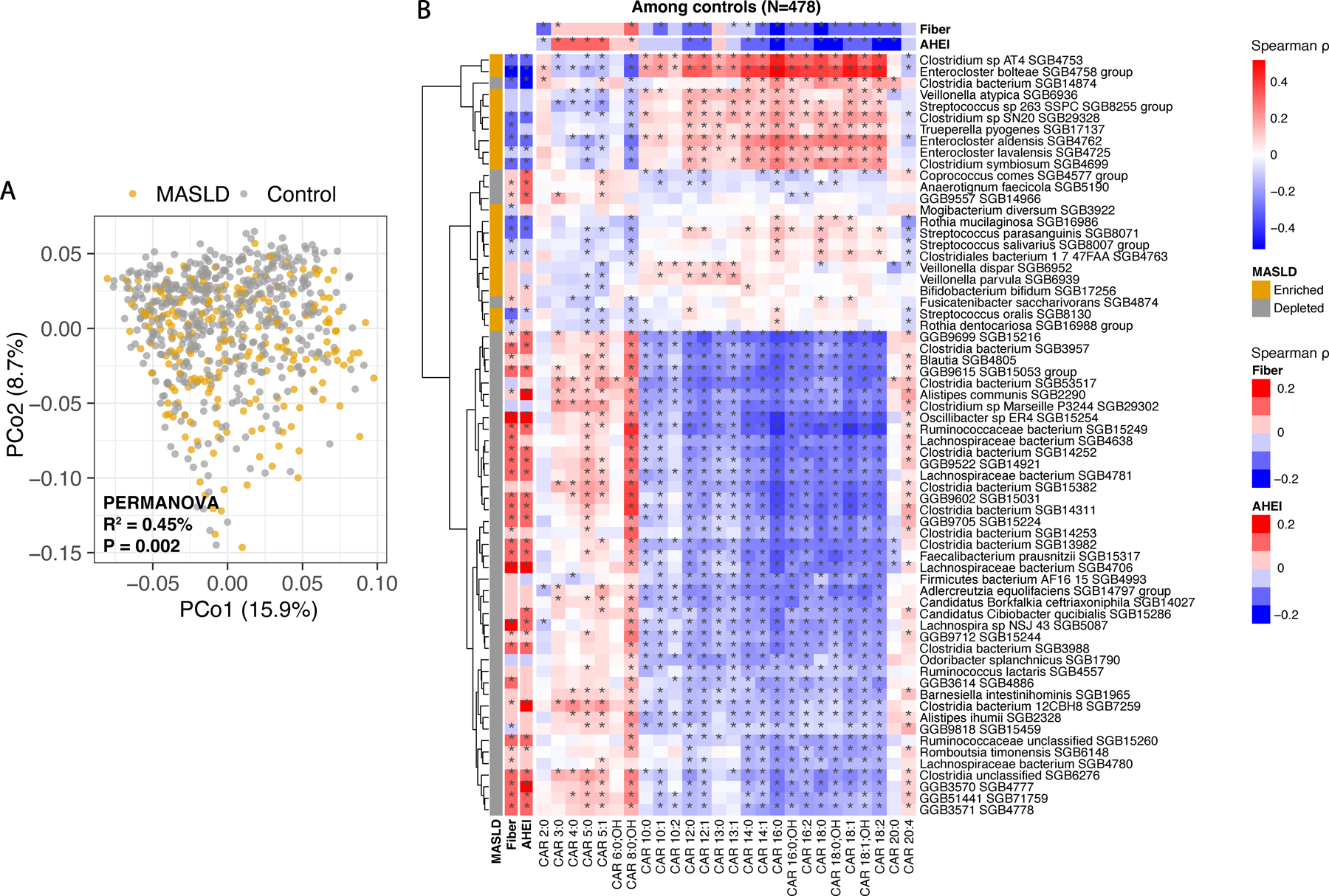
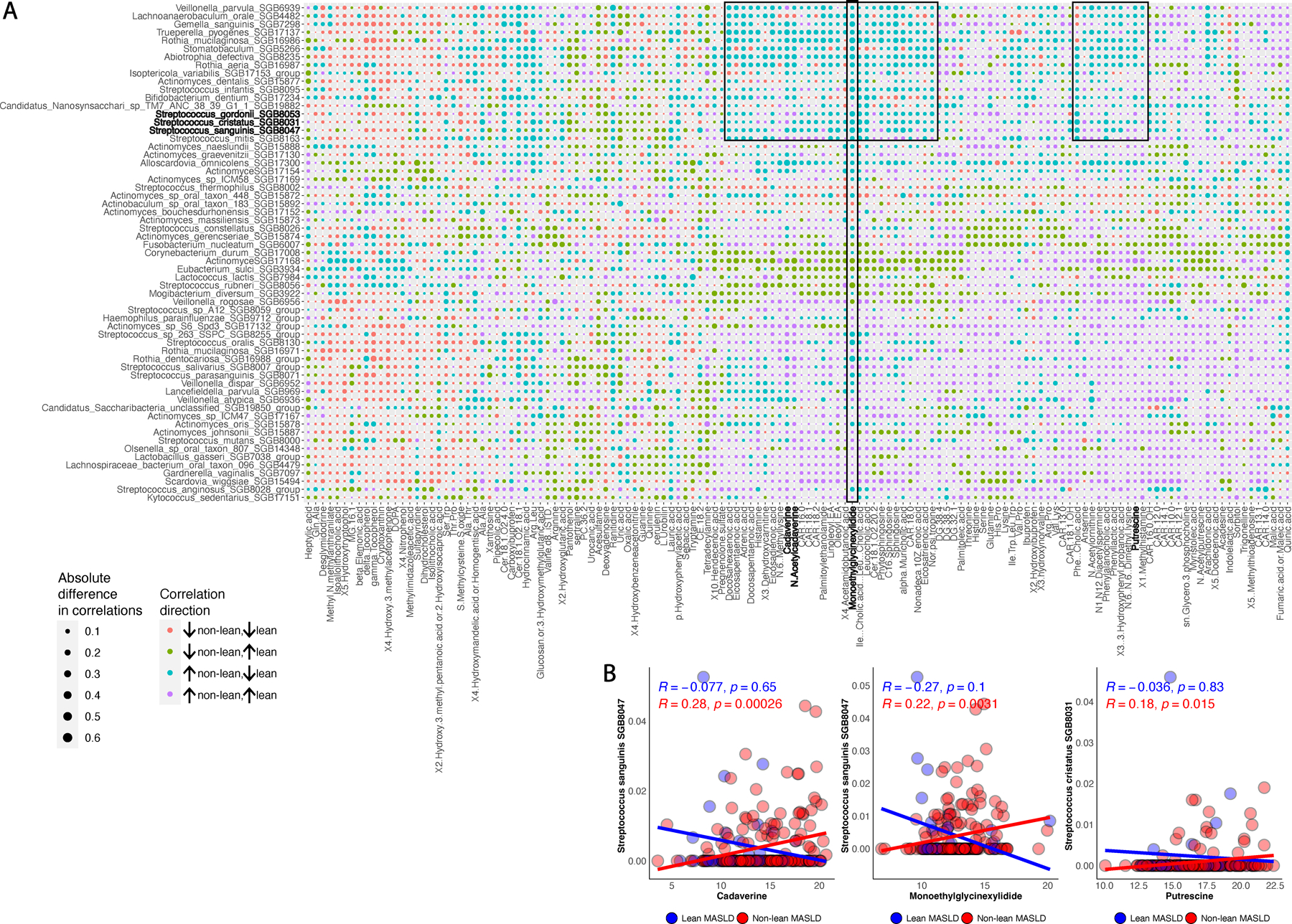
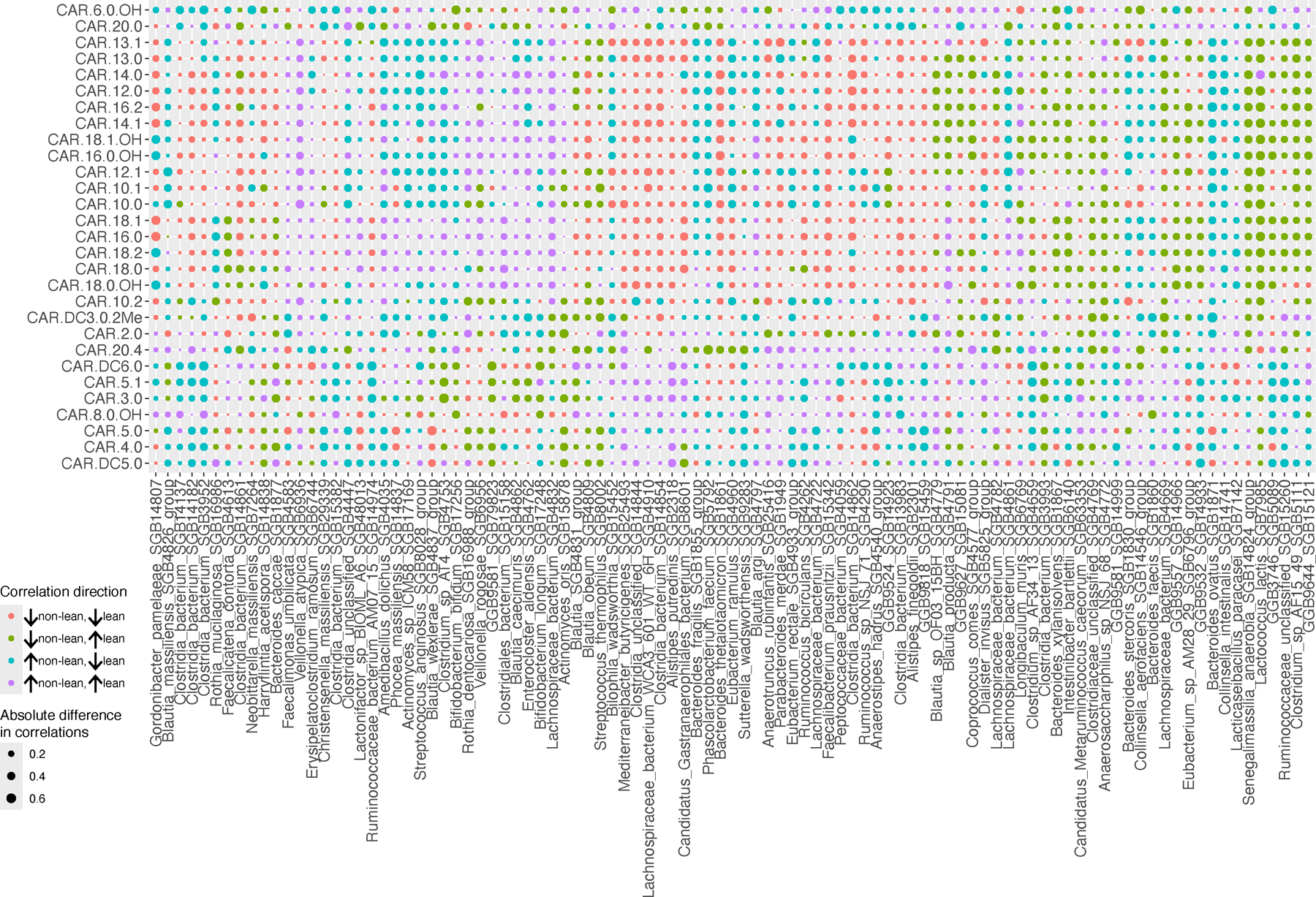
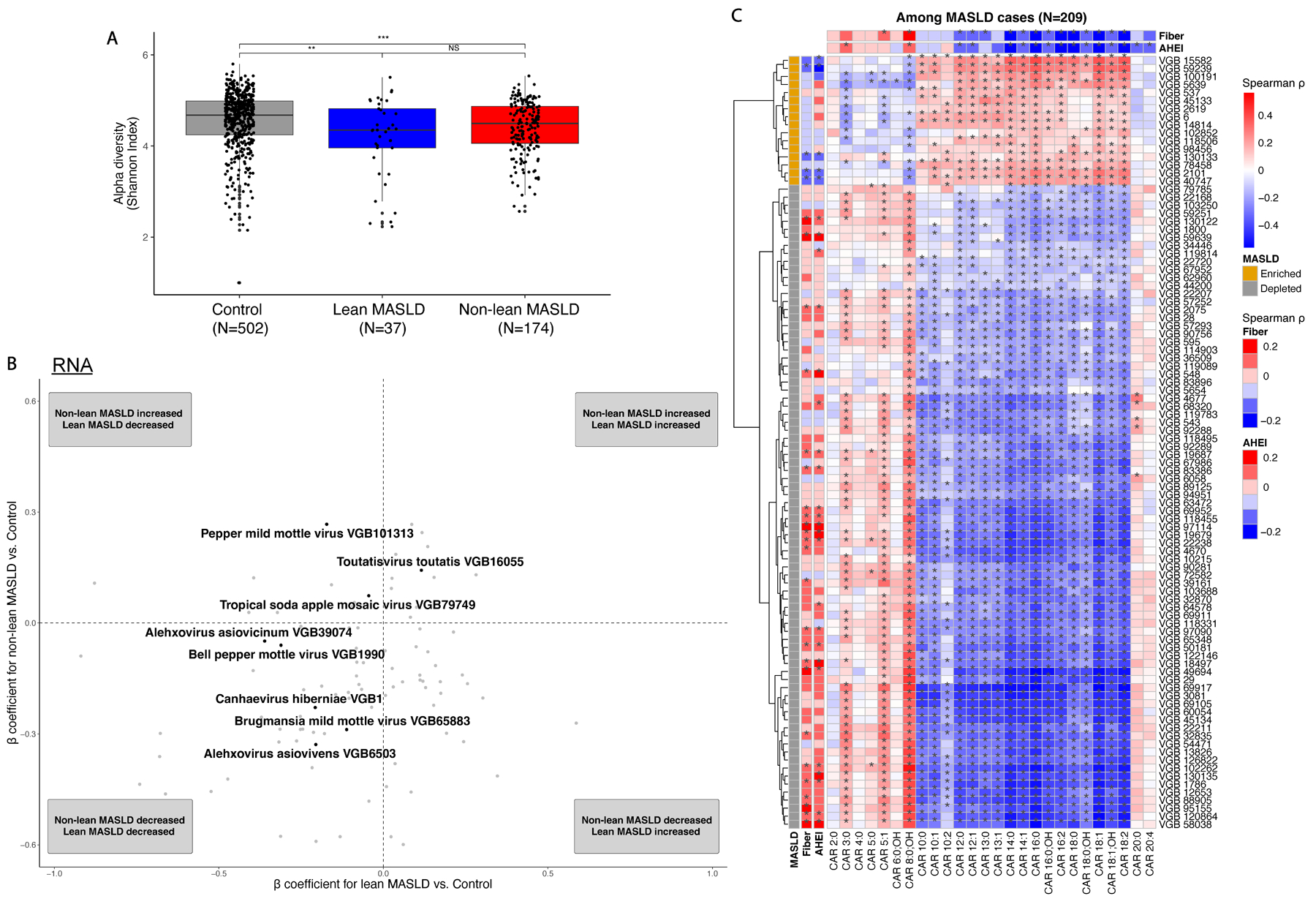
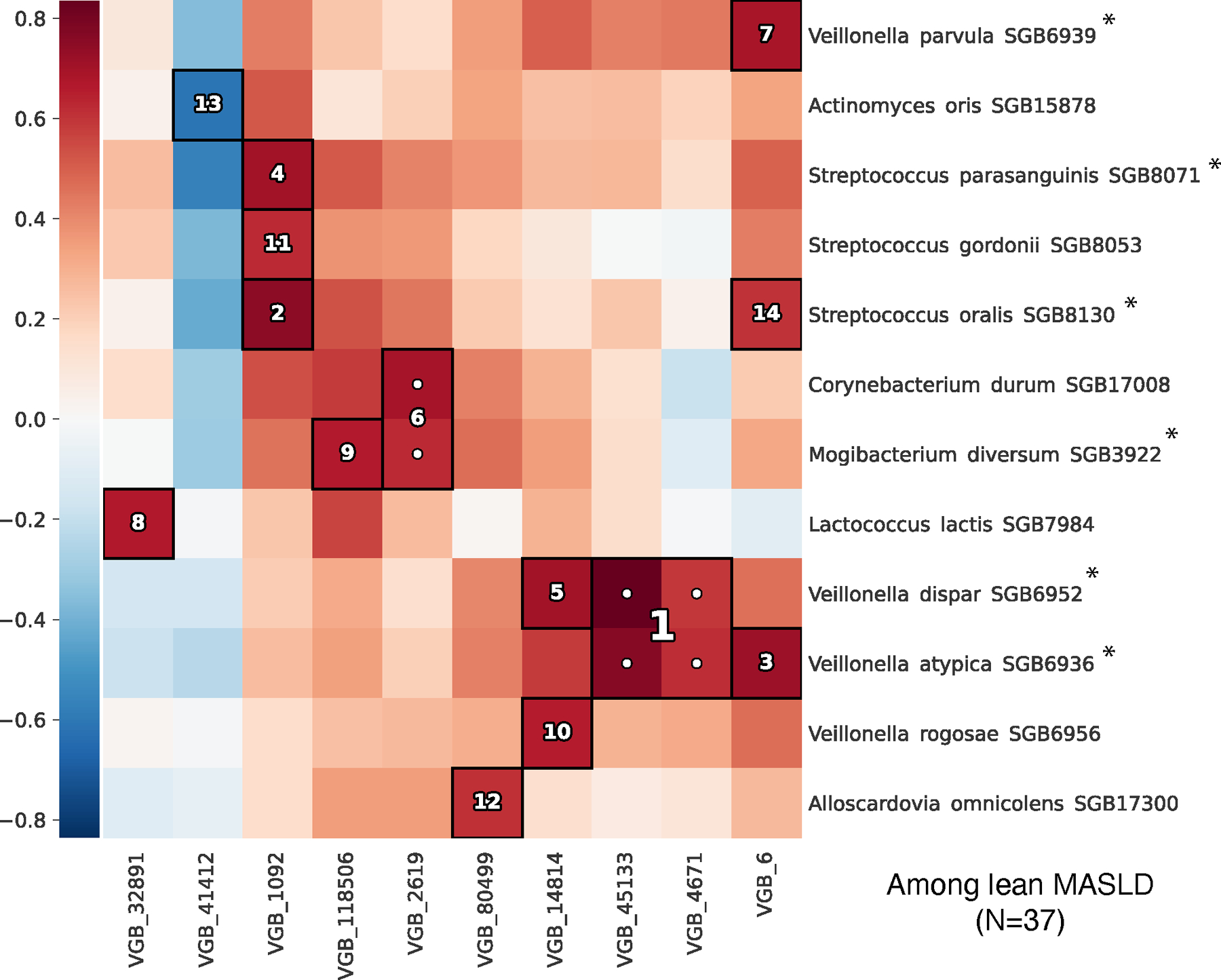
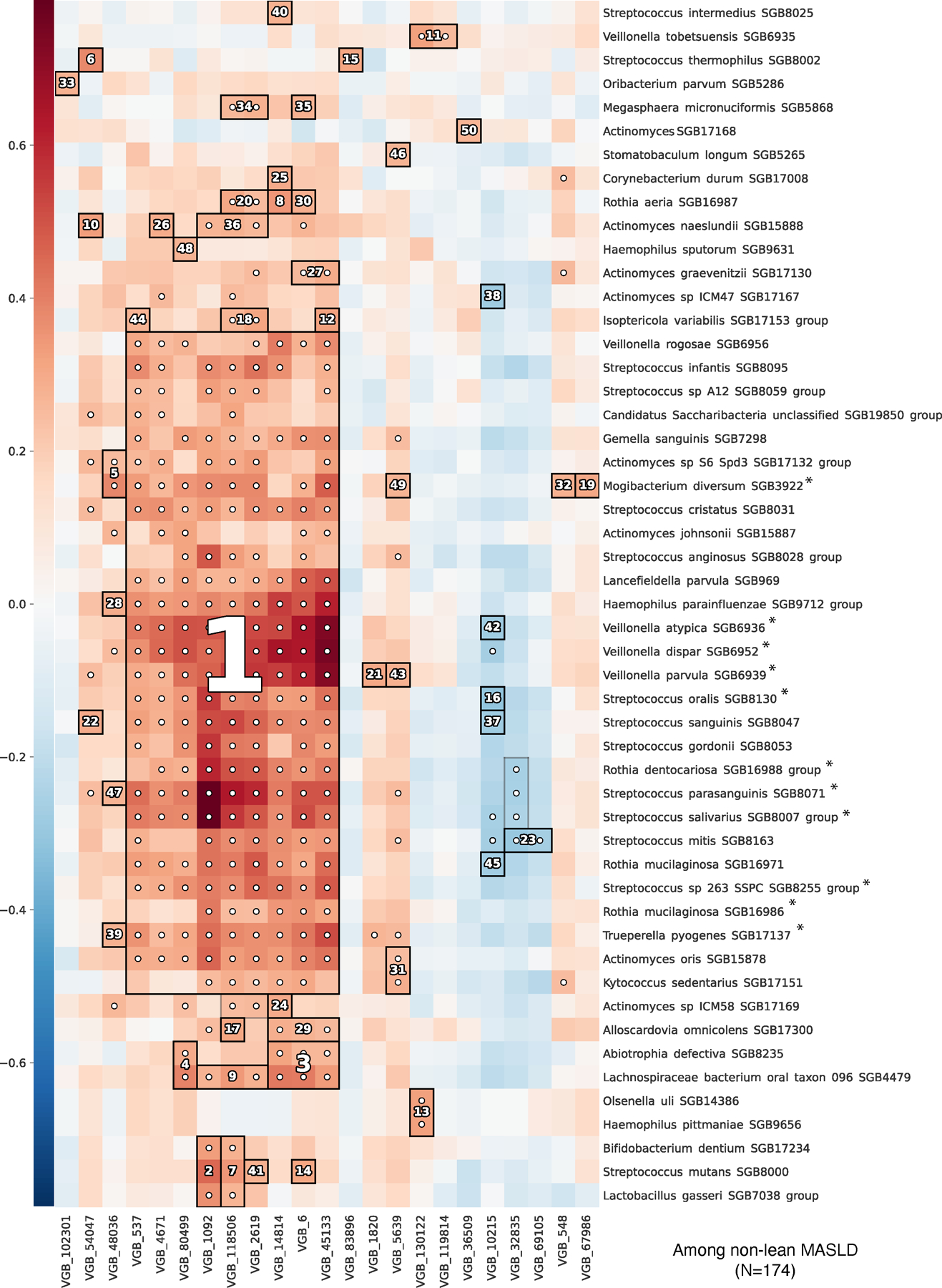
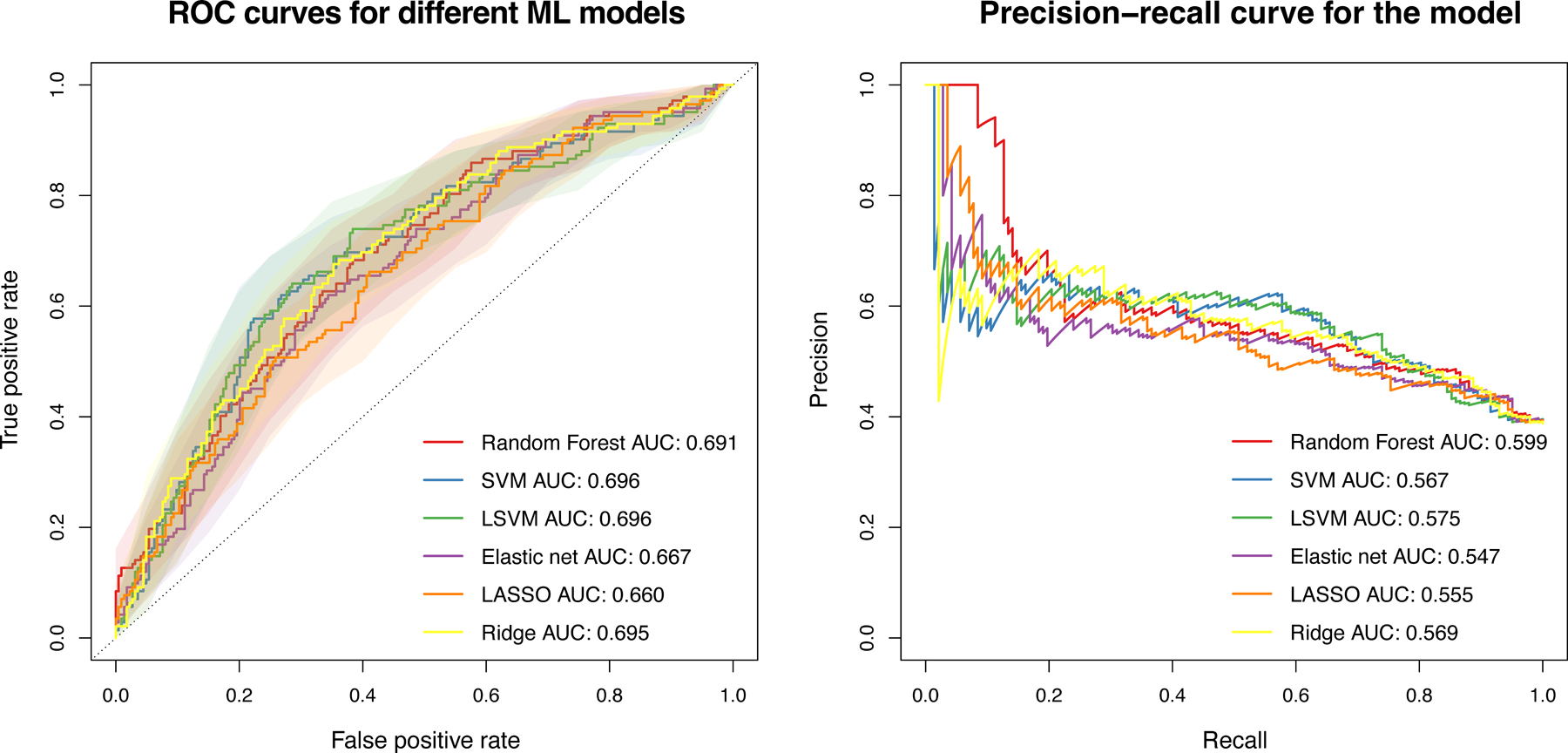
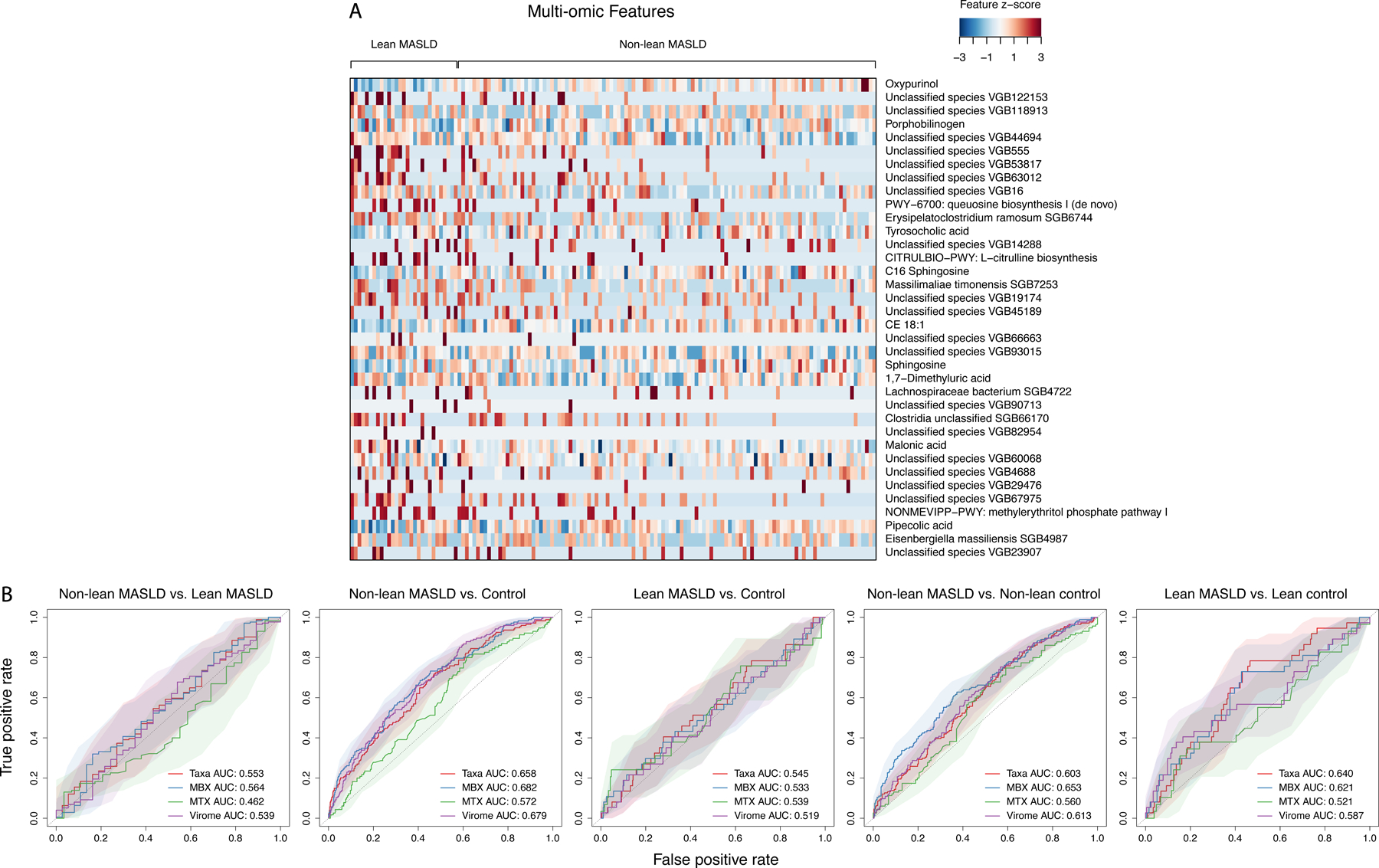
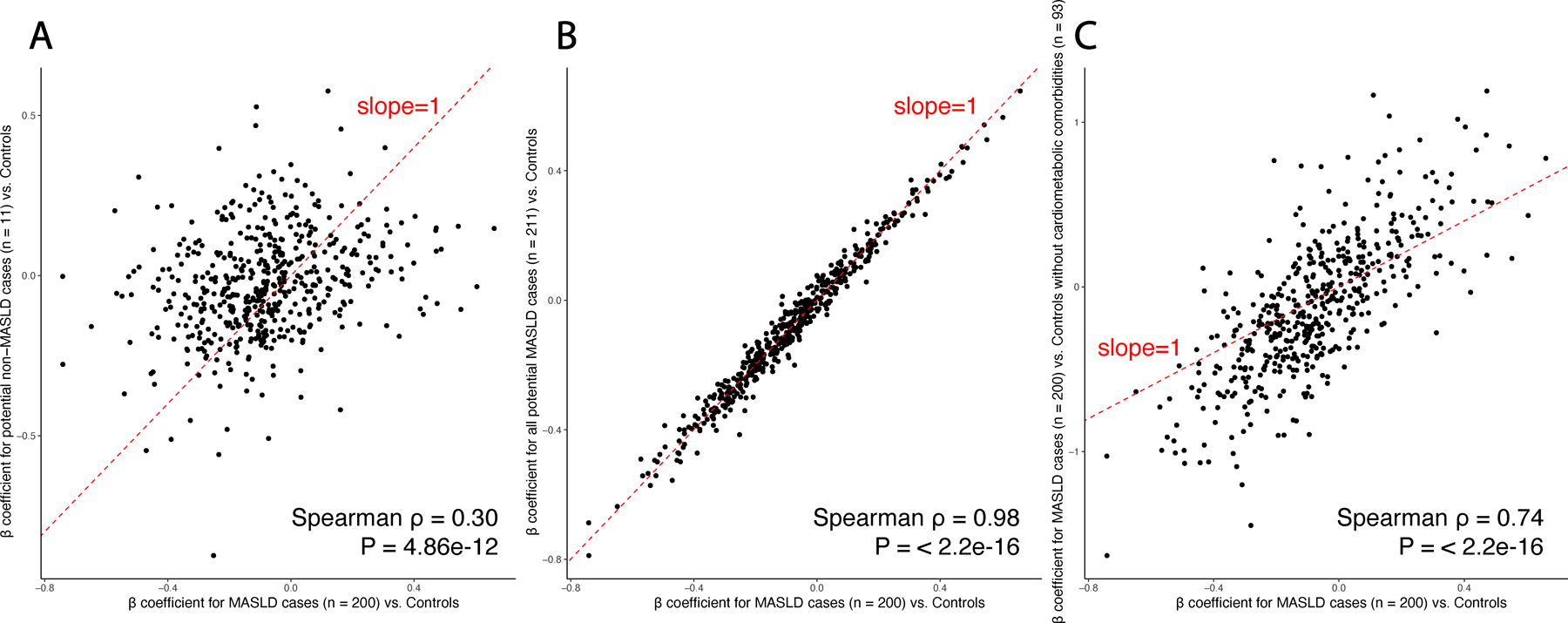
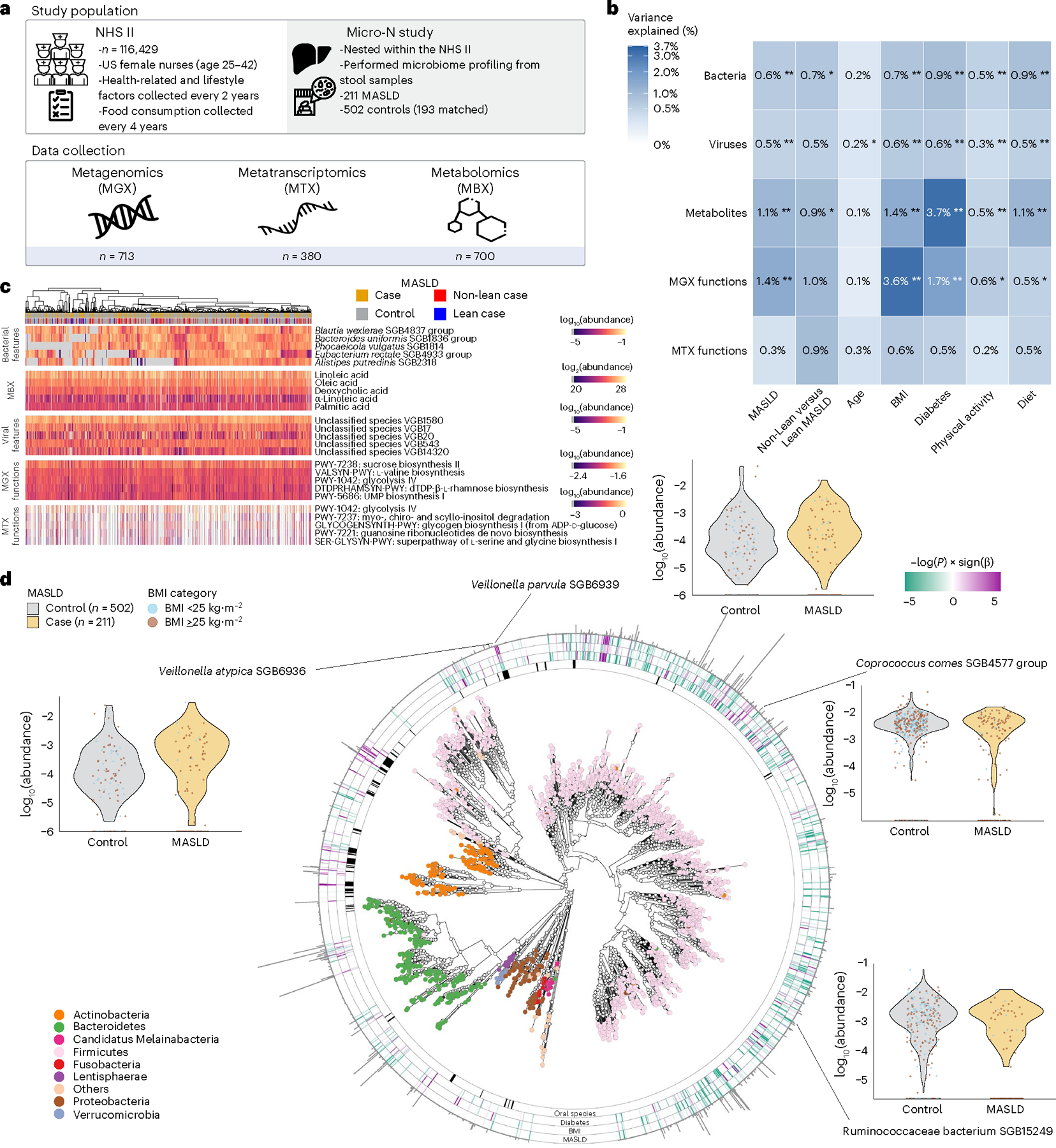
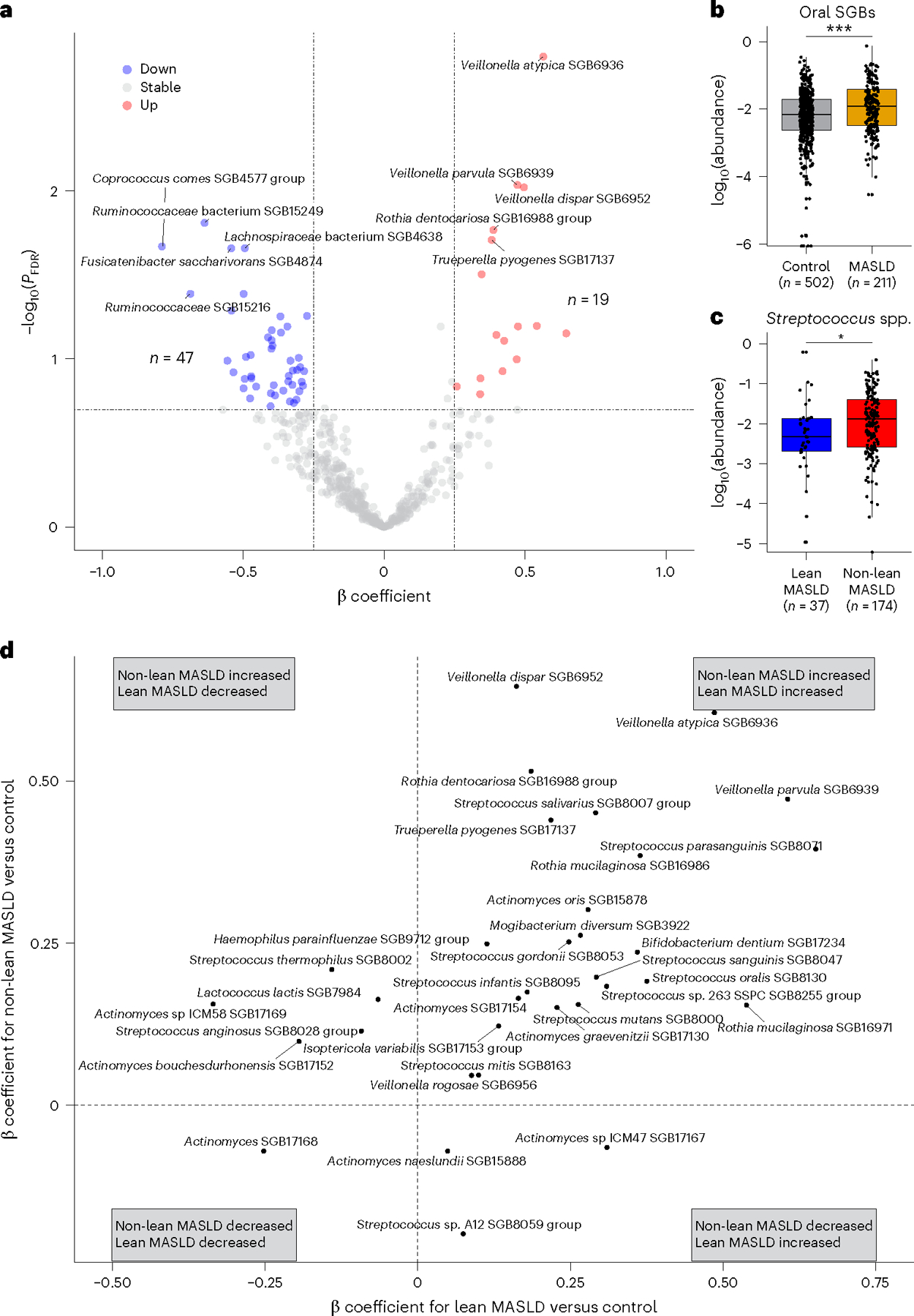
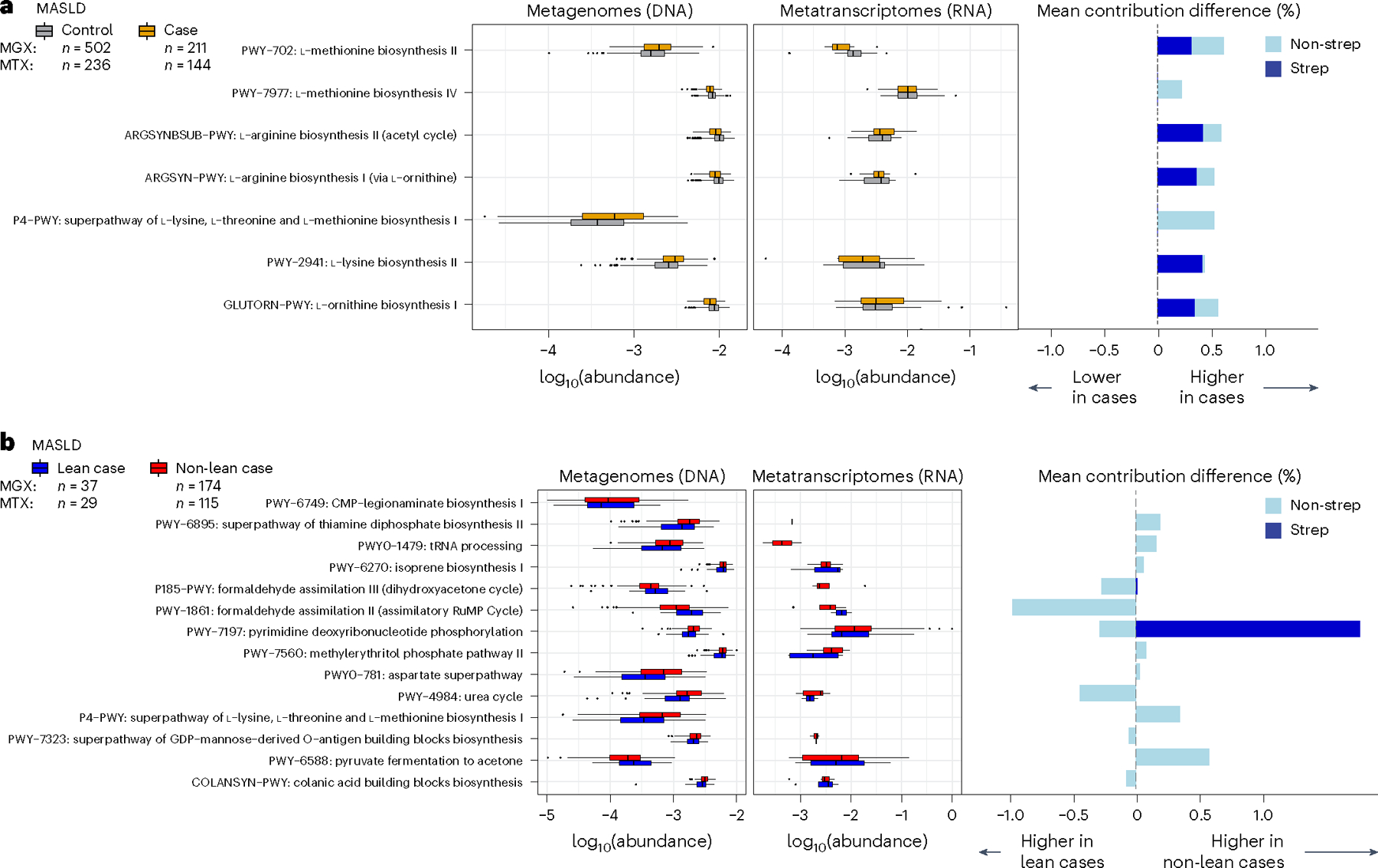
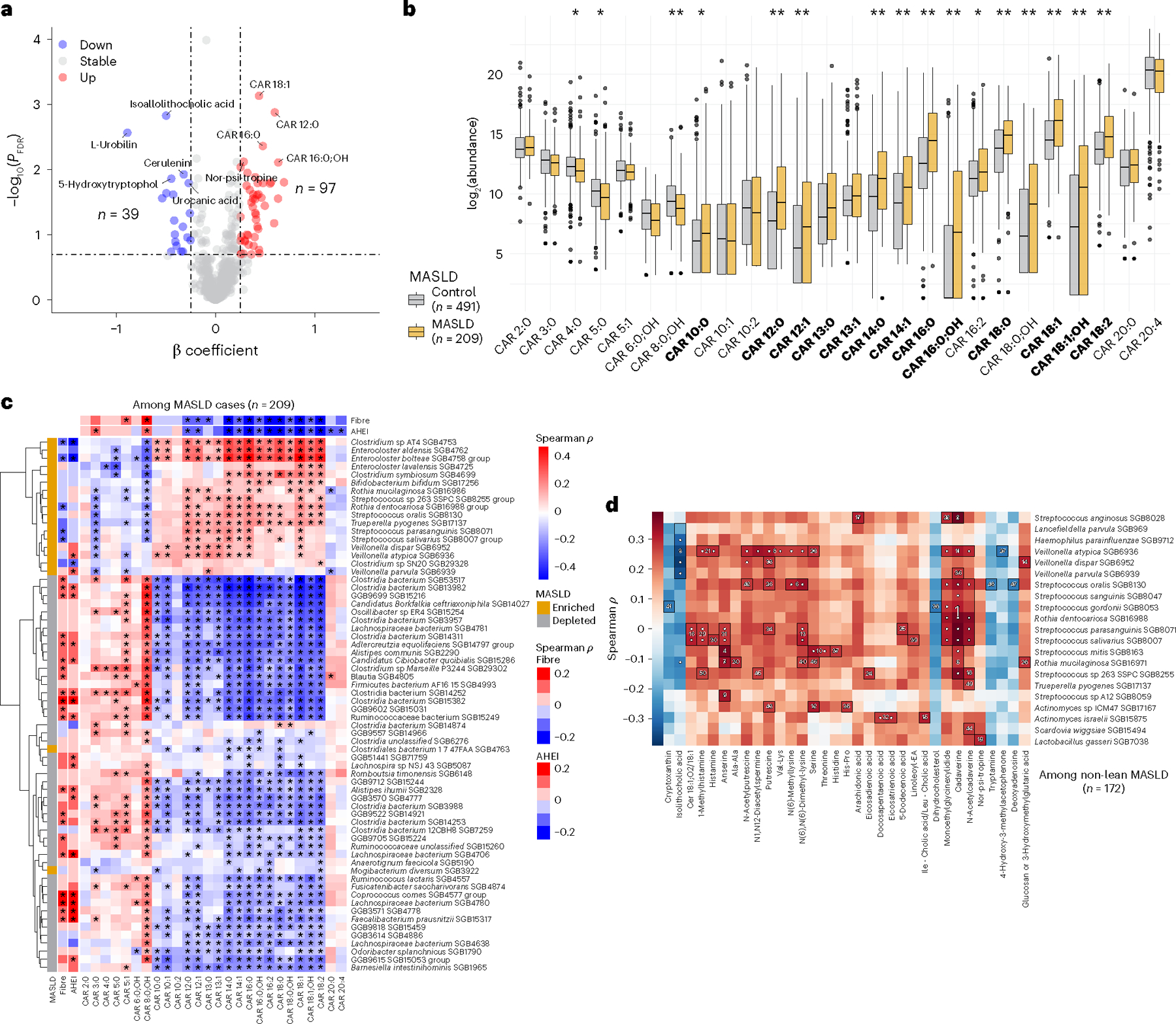
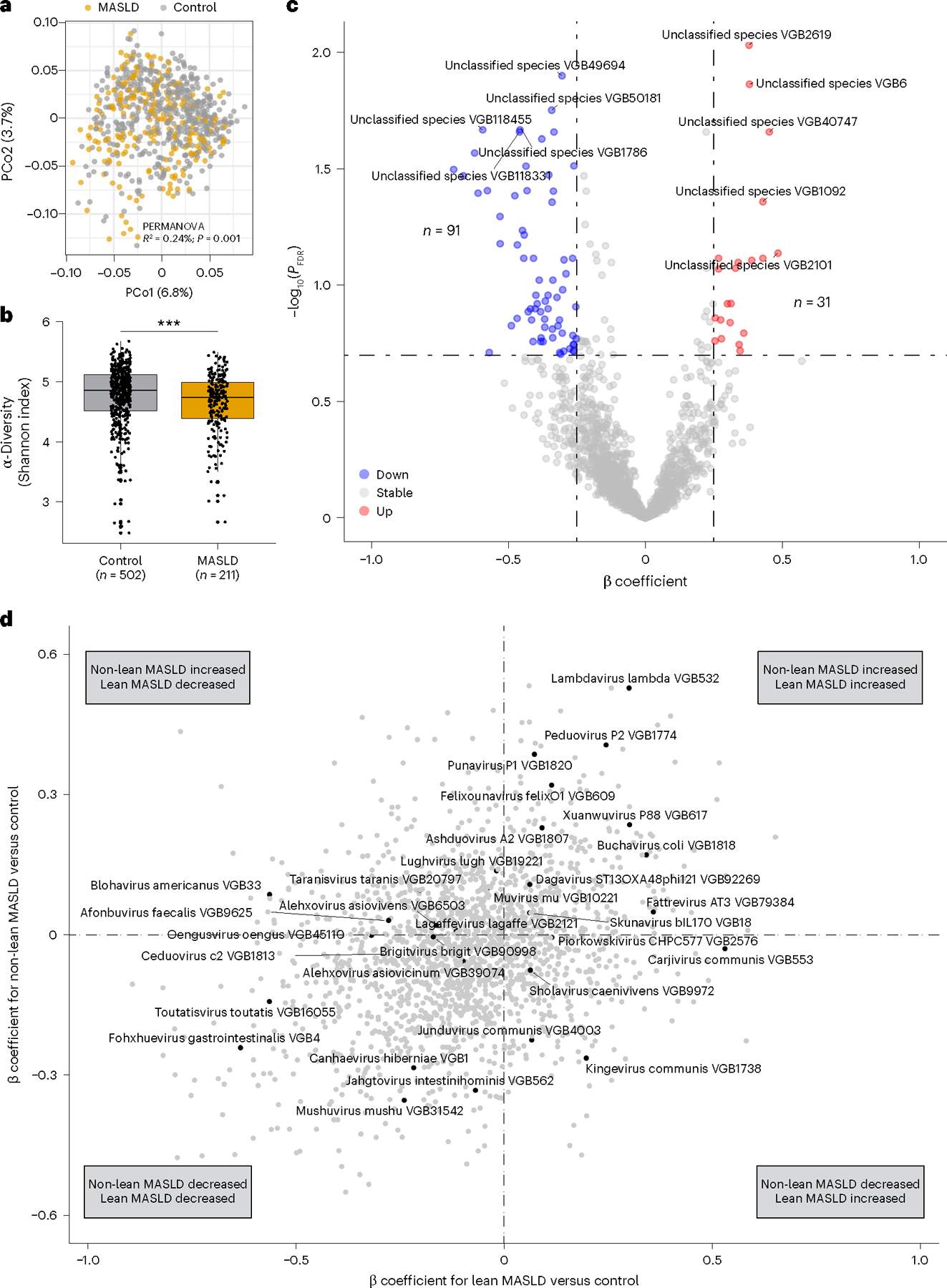
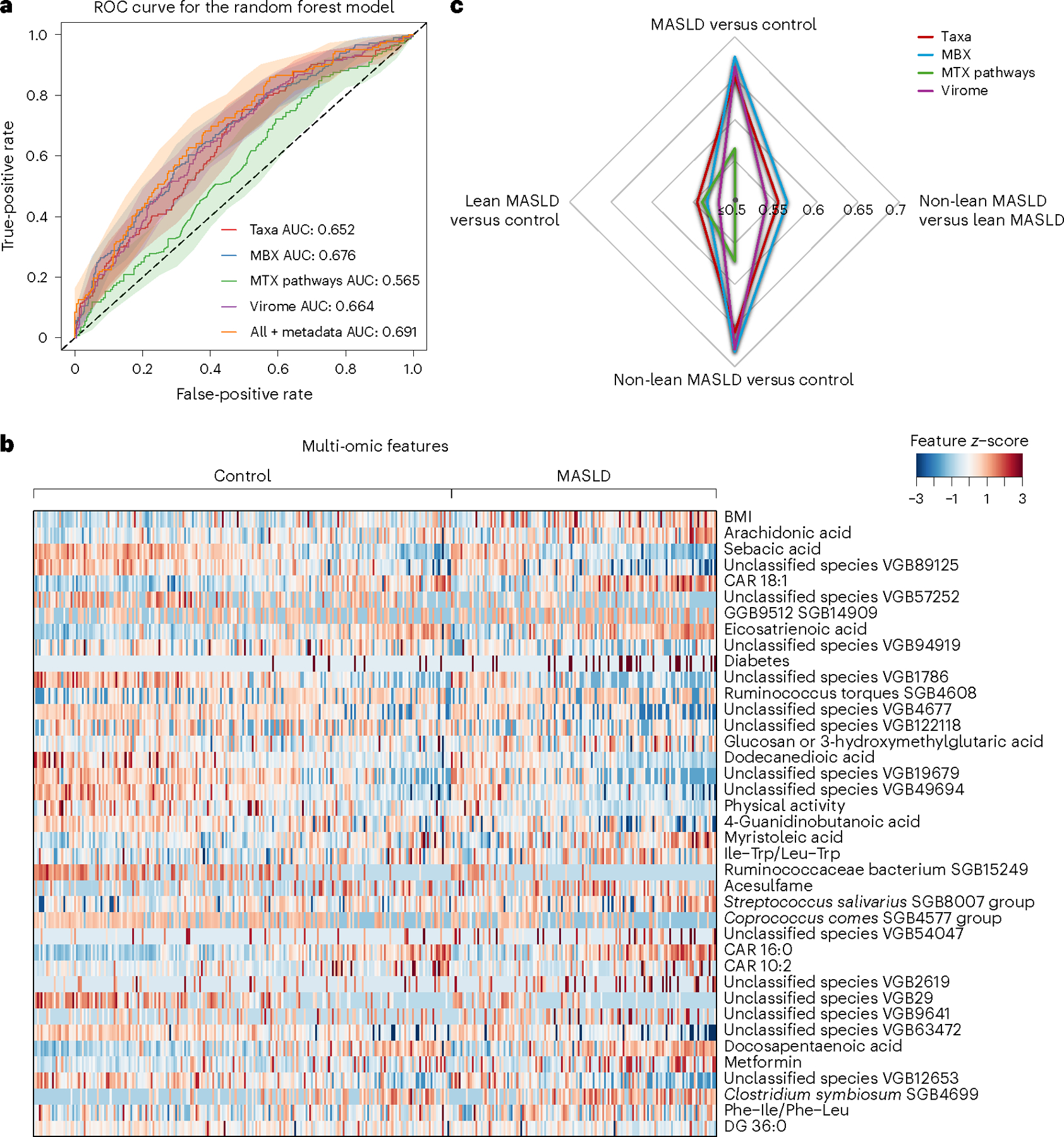
Metabolic dysfunction-associated steatotic liver disease (MASLD) is a common condition linked to obesity and the metabolic syndrome, yet its transkingdom connections have been under-investigated. We performed high-resolution multi-omic profiling-including stool metagenomes, metatranscriptomes and metabolomes-in 211 MASLD cases and 502 controls from a cohort of female nurses. Here we show that MASLD is associated with shifts in 66 gut bacterial species, including widespread enrichment of oral-typical microbes, and transkingdom dysbiosis involving not only bacterial but also viral taxa. Streptococcus spp. are more abundant in non-lean versus lean MASLD, the latter being a paradoxical subtype of a disease typically associated with increased adiposity. These microbial changes correspond with shifts in transcripts and metabolites, including increases in polyamines and acylcarnitines and reductions in secondary bile acids. We highlight gut viral perturbations in MASLD, showing that expansions of bacteriophage targeting oral-typical bacteria correspond to expansions of their bacterial hosts in the gut. We provide a comprehensive resource for understanding MASLD and highlight transkingdom multi-omic microbial shifts as potential contributors to its aetiopathogenesis.
© 2025. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: P.N. and B.H. were employees of Empress Therapeutics. C.H. is on the Scientific Advisory Board of Empress Therapeutics, Seres Therapeutics and ZOE Nutrition. All others declare no competing interests.
Figures
References
MeSH terms
Grants and funding
- R24DK110499/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R35CA253185/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- U01 CA176726/CA/NCI NIH HHS/United States
- Research Professorship/American Cancer Society (American Cancer Society, Inc.)
- K23DK125838/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- K23 DK125838/DK/NIDDK NIH HHS/United States
- Career Development Award/Crohn's and Colitis Foundation (Crohn's & Colitis Foundation)
- T32 CA009001/CA/NCI NIH HHS/United States
- R35 CA253185/CA/NCI NIH HHS/United States
- T32CA009001/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R24 DK110499/DK/NIDDK NIH HHS/United States
- Research Scholars Award/American Gastroenterological Association (AGA)
LinkOut - more resources
Full Text Sources
Miscellaneous
