

The Interplay of Ontogeny and Phylogeny at the Transcriptome Level of the Tetrapod Heart
- PMID: 40605499
- PMCID: PMC12328841
- DOI: 10.1002/jez.b.23312
The Interplay of Ontogeny and Phylogeny at the Transcriptome Level of the Tetrapod Heart
Abstract
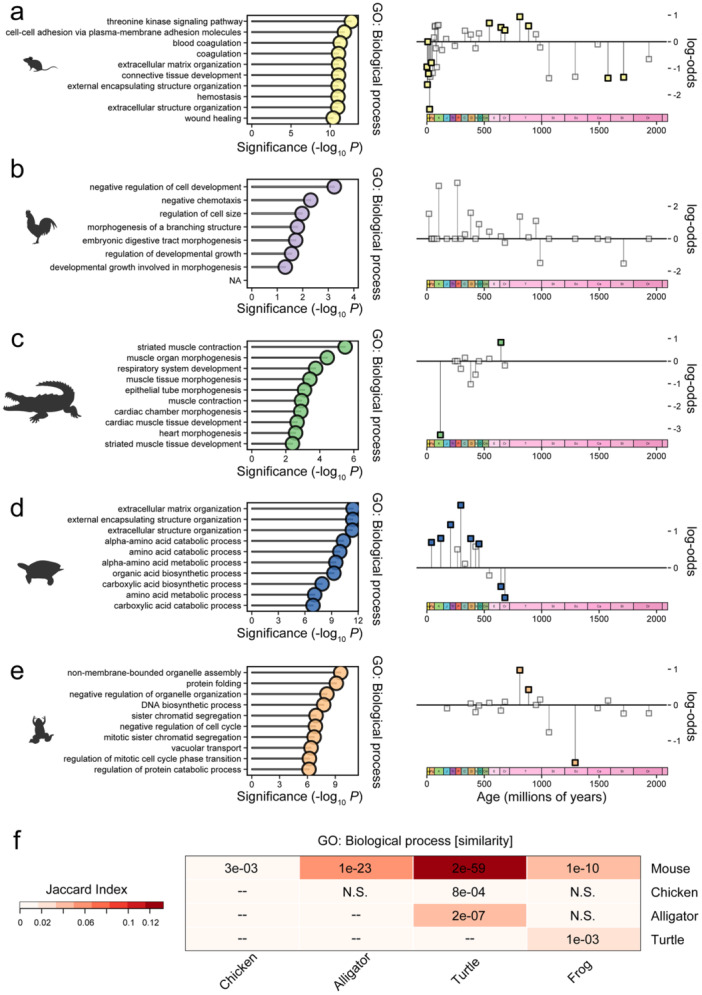
The tetrapod heart is characterized by three chambers in amphibians and non-avian reptiles, as opposed to four in birds, crocodilians and mammals. We explored this diversity via the most phylogenetically comprehensive comparison of heart transcriptomes undertaken to date. Transcriptomes representing the ontogeny of heart compartmentalization (septation) in alligator, chicken, frog, mouse, lizard and turtle embryos exhibited a clear species-specific signal, which was driven by genes involved in heart contraction. During the stage dominated by septation-related tissue transformations, the most highly expressed genes shared by species originated before the tetrapods diversified and were related to septum morphogenesis, ventricular development, and chamber formation. The expression of septation-related genes did not adhere to phylogeny or heart chamber number, and genes differentially expressed across developmental stages within species varied in their evolutionary ages and predicted functions. We discuss how the acquisition of novel structures in some lineages, convergent evolution of four heart chambers, embryonic metabolism, microstructural variation, and ontogenetic shifts (heterochronies), collectively, provide insight into evolved and conserved patterns of transcriptome-level variation. These data serve as a resource to further stimulate evo-devo research on complex organ systems, such as the heart.
Keywords: comparative transcriptomics; evo‐devo; heart development; tetrapod embryology.
© 2025 The Author(s). Journal of Experimental Zoology Part B: Molecular and Developmental Evolution published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





Similar articles
-
Embryonic hypoxia alters cardiac gene expression patterns in American alligators, Alligator mississippiensis.Physiol Genomics. 2025 Sep 1;57(9):499-517. doi: 10.1152/physiolgenomics.00192.2024. Epub 2025 Jun 25. Physiol Genomics. 2025. PMID: 40560627
-
Stage-specific DNA methylation dynamics in mammalian heart development.Epigenomics. 2025 Apr;17(5):359-371. doi: 10.1080/17501911.2025.2467024. Epub 2025 Feb 21. Epigenomics. 2025. PMID: 39980349 Review.
-
A history of normal plates, tables and stages in vertebrate embryology.Int J Dev Biol. 2007;51(1):1-26. doi: 10.1387/ijdb.062189nh. Int J Dev Biol. 2007. PMID: 17183461 Free PMC article.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
Genomic and transcriptomic insights into vertebrate host-specific Lactobacillus johnsonii adaptation in the gastrointestinal tract.mSphere. 2025 Jun 25;10(6):e0005225. doi: 10.1128/msphere.00052-25. Epub 2025 May 13. mSphere. 2025. PMID: 40358235 Free PMC article.
References
-
- von Baer, K. E. (1828). Über Entwickelungsgeschichte der Thiere: Beobachtung und Reflexion (Vol. 1): Bornträger.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
