

Species aggregation models resolve essential foraging habitat: Implications for conservation and management
- PMID: 40605787
- PMCID: PMC12223939
- DOI: 10.1002/eap.70068
Species aggregation models resolve essential foraging habitat: Implications for conservation and management
Abstract
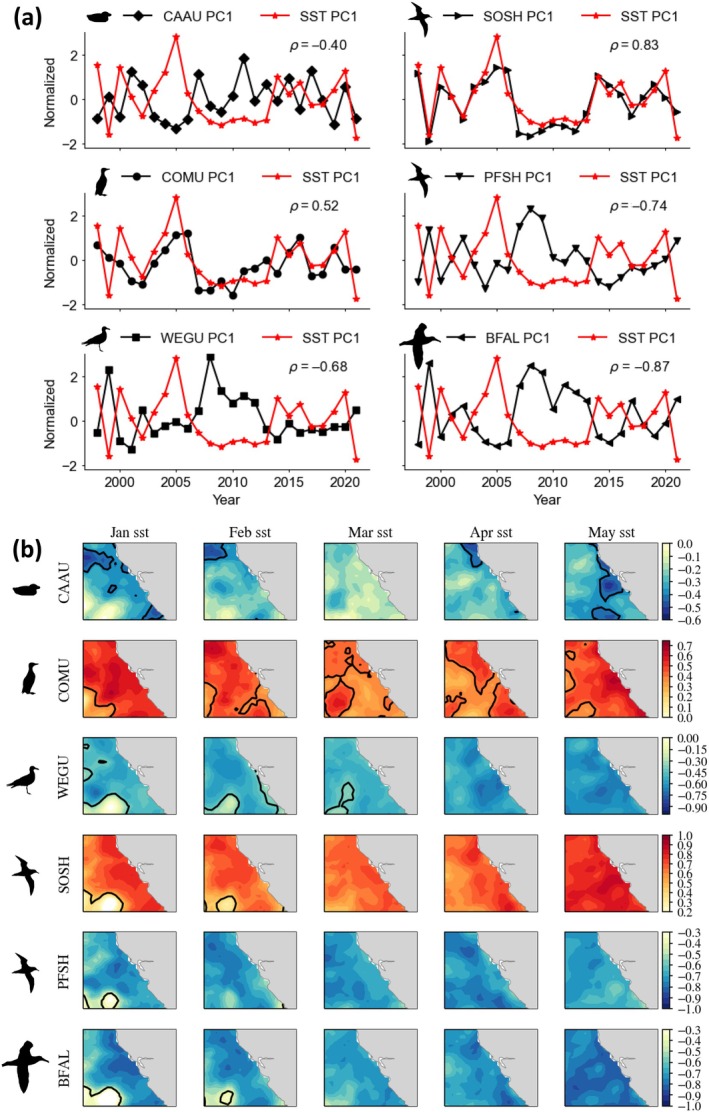
Species aggregations are a culmination of behavioral events arising from an array of biophysical interactions, dynamically shifting in space and time. Prediction of species' aggregation dynamics remains a challenge in studies of their distribution patterns. Species distribution models (SDMs) are statistical tools for understanding spatial patterns of marine biodiversity, ranging from essential species habitat, assessing fisheries bycatch, and projecting future distribution changes. SDMs involving pelagic species abundance generally do not typically resolve aggregation patterns. We use a 20-year observation record of seabird species aggregations, with seabirds being the most easily quantified "pelagic" species, to develop SDMs and a regional ocean modeling system to identify physical drivers and changes in aggregation location and intensity over time. We apply a conceptual ecosystem model to organize environmental covariates according to habitat production within coastal upwelling systems. The SDM used a 2-step modeling approach: a presence/absence model and a binary aggregation model. Thus, we aim to predict factors that characterize baseline ocean habitat for a species (presence/absence) and that aggregate large numbers of the species. Prediction of seabird aggregation results in realistic spatial distribution patterns that reflect known species habitat associations. Temporally, aggregation indices indicate mixed responses both within and between resident and migrant species, reflecting interannual effects of warm/cool ocean years and mesoscale structure supporting enhanced or decreased productive foraging habitat. The most abundant species were more likely to form aggregations during warmer years, indicating a response to a decrease in productive foraging habitat. The occurrence of species aggregations in spring is predictable by examining ocean-climate conditions in the preceding winter, thus providing a potential early warning system of anticipated ecosystem shifts. We contend that the aggregation occurrence model may improve the realism of pelagic SDMs and their utility for assessing spatial and temporal variability of trophic interactions. We discuss the utility of species aggregation models for quantifying the variability in critical pelagic habitats, the ecology and response of seabird species as indicators, advancement of ecosystem modeling and monitoring, and conservation applications (e.g., bycatch, wind energy, and oil spills).
Keywords: aggregation; conservation; distribution; ecosystem oceanography; foraging habitat; patchiness; realism; spatial organization; species distribution model.
© 2025 The Author(s). Ecological Applications published by Wiley Periodicals LLC on behalf of The Ecological Society of America.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







Similar articles
-
The potential for species distribution models to distinguish source populations from sinks.J Anim Ecol. 2024 Dec;93(12):1924-1934. doi: 10.1111/1365-2656.14201. Epub 2024 Oct 21. J Anim Ecol. 2024. PMID: 39429222
-
Temperature sensitivity of the interspecific interaction strength of coastal marine fish communities.Elife. 2023 Jul 11;12:RP85795. doi: 10.7554/eLife.85795. Elife. 2023. PMID: 37431235 Free PMC article.
-
Space-for-time substitutions exaggerate urban bird-habitat ecological relationships.J Anim Ecol. 2024 Dec;93(12):1854-1867. doi: 10.1111/1365-2656.14194. Epub 2024 Nov 6. J Anim Ecol. 2024. PMID: 39506196 Free PMC article.
-
A scoping review of tropical pioneer trees' roles for restoration and conservation management: Harungana madagascariensis (Hypericaceae) a widespread African species as a model.PeerJ. 2025 May 23;13:e19458. doi: 10.7717/peerj.19458. eCollection 2025. PeerJ. 2025. PMID: 40421372 Free PMC article.
-
What is the evidence documenting the effects of marine or coastal nature conservation or natural resource management activities on human well-being in South East Asia? A systematic map.Environ Int. 2021 Jun;151:106397. doi: 10.1016/j.envint.2021.106397. Epub 2021 Mar 10. Environ Int. 2021. PMID: 33713939
References
-
- Adams, J. , MacLeod C., Suryan R. M., Hyrenbach K. D., and Harvey J. T.. 2012. “Summer‐Time Use of West Coast U.S. National Marine Sanctuaries by Migrating Sooty Shearwaters (Puffinus griseus).” Biological Conservation 156: 105–116.
-
- Ainley, D. G. , and Boekelheide R. J., eds. 1990. Seabirds of the Farallon Islands: Ecology, Dynamics, and Structure of an Upwelling‐System Community 450. Stanford, CA: Stanford University Press.
-
- Ainley, D. G. , DeGange A. R., Jones L. L., and Beach R. J.. 1981. “Mortality of Seabirds in High‐Seas Salmon Gill Nets.” Fishery Bulletin 79: 800–806.
-
- Ainley, D. G. , Dugger K. D., Ford R. G., Pierce S. D., Reese D. C., Brodeur R. D., Tynan C. T., and Barth J. A.. 2009. “The Spatial Association of Predators and Prey at Frontal Features in the Northern California Current: Competition, Facilitation, or Merely Co‐Occurrence?” Marine Ecology Progress Series 389: 271–294.
-
- Ainley, D. G. , and Hyrenbach K. D.. 2010. “Top‐Down and Bottom‐up Factors Affecting Seabird Population Trends in the California Current System (1985–2006).” Progress in Oceanography 84: 242–254. 10.1016/j.pocean.2009.10.001. - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
