

Plasma membrane remodeling in GM2 gangliosidoses drives synaptic dysfunction
- PMID: 40608809
- PMCID: PMC12251256
- DOI: 10.1371/journal.pbio.3003265
Plasma membrane remodeling in GM2 gangliosidoses drives synaptic dysfunction
Abstract
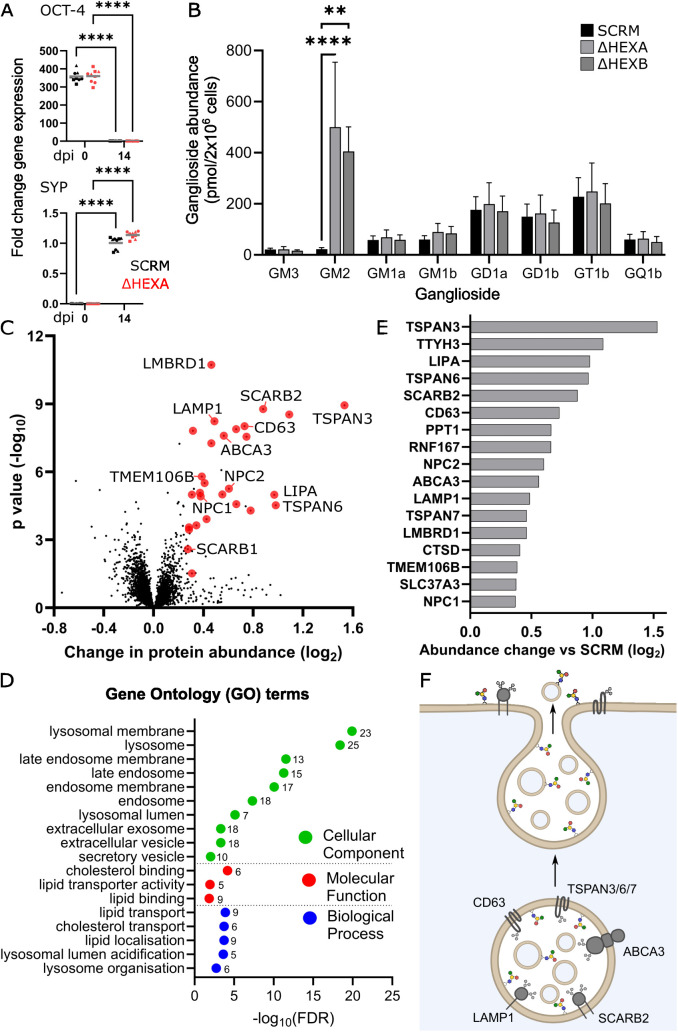
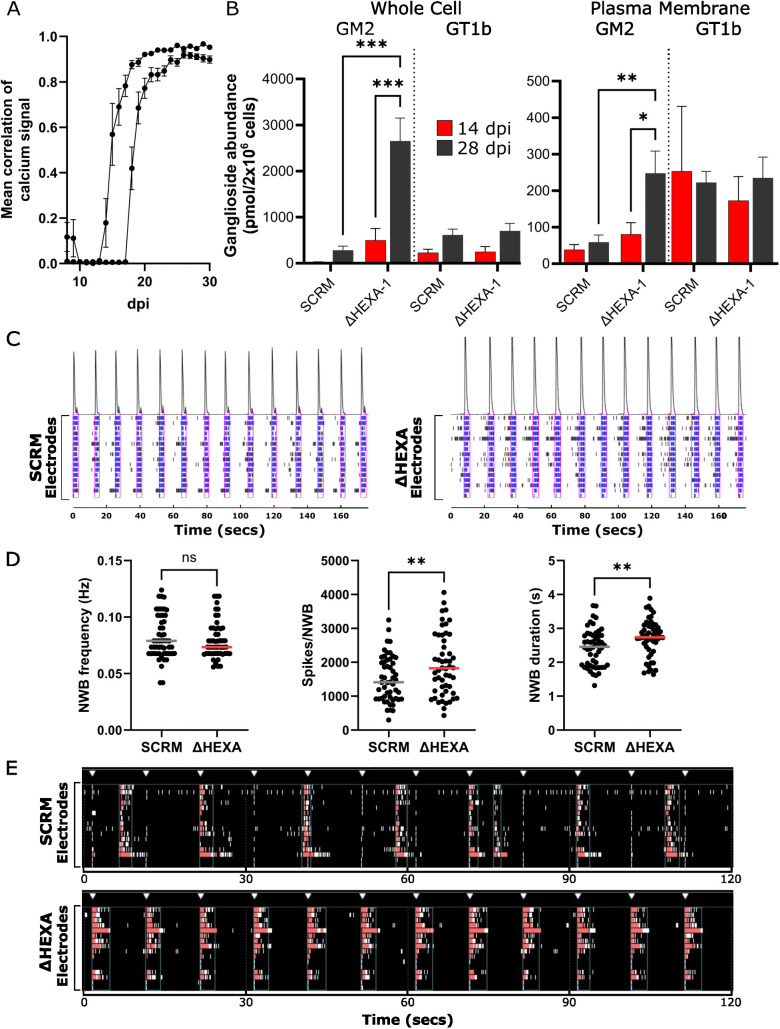
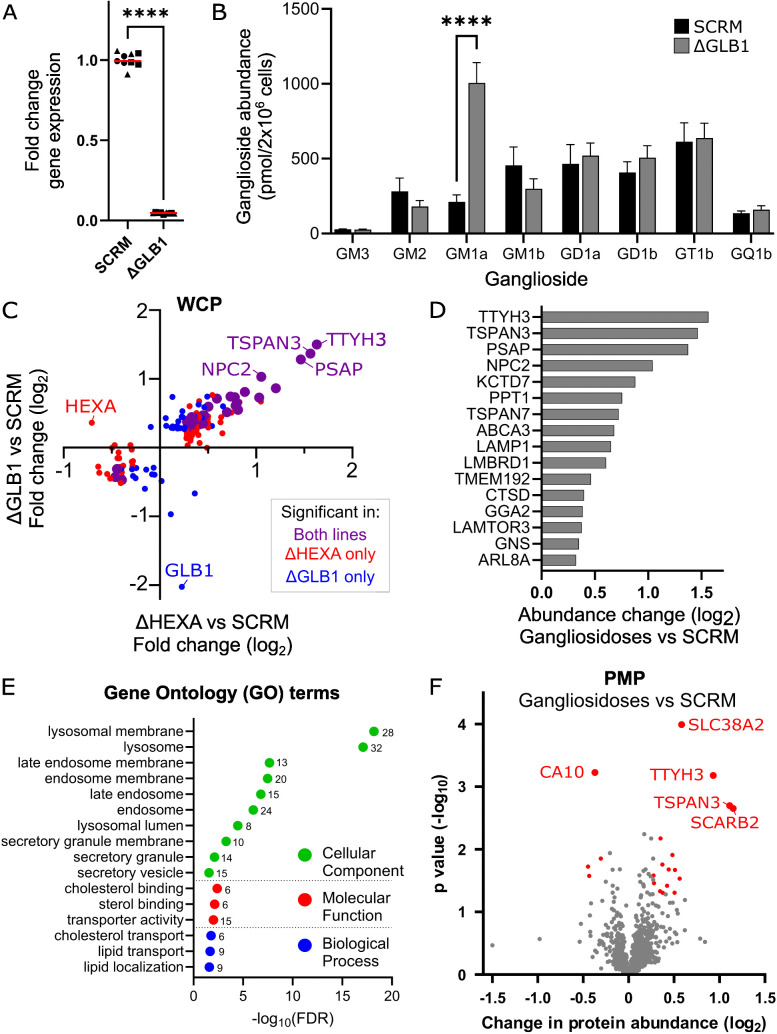
Glycosphingolipids (GSL) are important bioactive membrane components. GSLs containing sialic acids, known as gangliosides, are highly abundant in the brain and diseases of ganglioside metabolism cause severe early-onset neurodegeneration. The ganglioside GM2 is processed by β-hexosaminidase A and when non-functional GM2 accumulates causing Tay-Sachs and Sandhoff diseases. We have developed i3Neuron-based disease models demonstrating storage of GM2 and severe endolysosomal dysfunction. Additionally, the plasma membrane (PM) is significantly altered in its lipid and protein composition. These changes are driven in part by lysosomal exocytosis causing inappropriate accumulation of lysosomal proteins on the cell surface. There are also significant changes in synaptic protein abundances with direct functional impact on neuronal activity. Lysosomal proteins are also enriched at the PM in GM1 gangliosidosis supporting that lysosomal exocytosis is a conserved mechanism of PM proteome change in these diseases. This work provides mechanistic insights into neuronal dysfunction in gangliosidoses highlighting that these are severe PM disorders with implications for other lysosomal and neurodegenerative diseases.
Copyright: © 2025 Nicholson et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
