

The intracellular bacterium Orientia tsutsugamushi uses the autotransporter ScaC to activate BICD adaptors for dynein-based motility
- PMID: 40610402
- PMCID: PMC12229497
- DOI: 10.1038/s41467-025-61105-5
The intracellular bacterium Orientia tsutsugamushi uses the autotransporter ScaC to activate BICD adaptors for dynein-based motility
Abstract
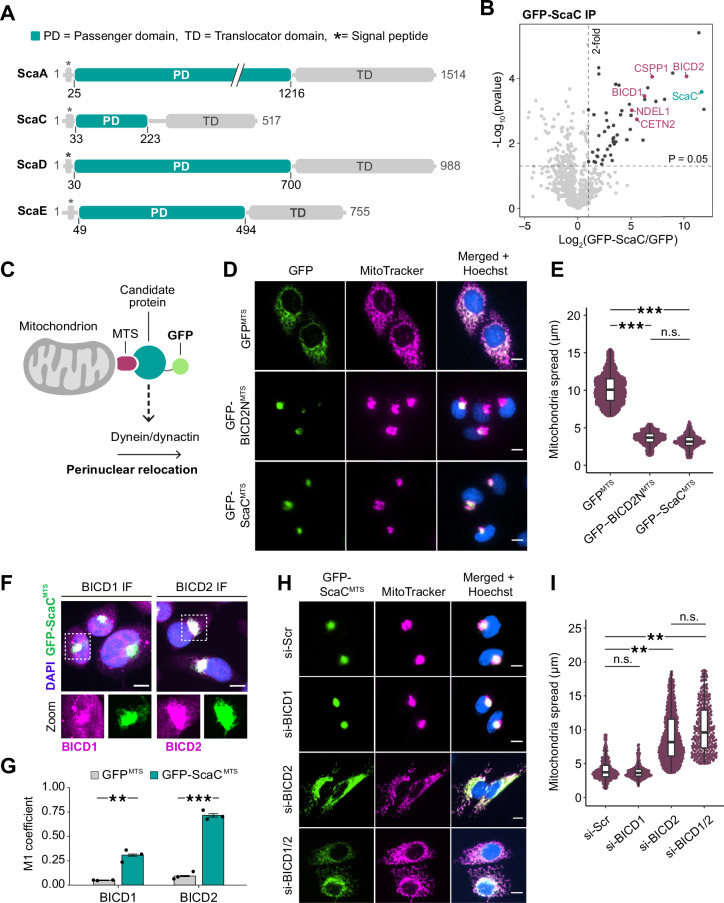
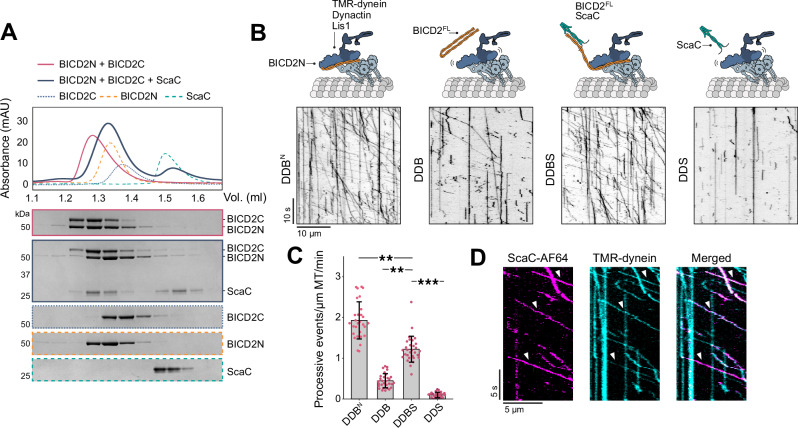
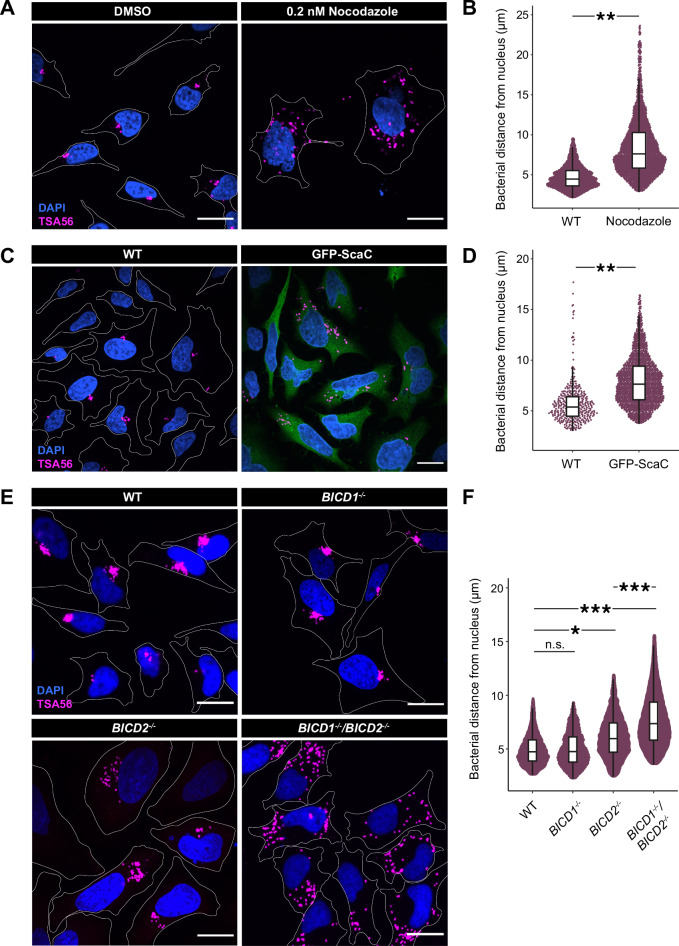
The intracellular bacterium Orientia tsutsugamushi relies on the microtubule cytoskeleton and the motor protein dynein to traffic to the perinuclear region within infected cells. However, it remains unclear how the bacterium is coupled to the dynein machinery and how transport is regulated. Here, we discover that O. tsutsugamushi uses its autotransporter protein ScaC to recruit the dynein adaptors BICD1 and BICD2 for movement to the perinucleus. We show that ScaC is sufficient to engage dynein-based motility in the absence of other bacterial proteins and that BICD1 and BICD2 are required for efficient movement of O. tsutsugamushi during infection. Using TIRF single-molecule assays, we demonstrate that ScaC induces BICD2 to adopt an open conformation which activates the assembly of dynein-dynactin complexes. Our results reveal a role for BICD adaptors during bacterial infection and provide mechanistic insights into the life cycle of an important human pathogen.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures


References
-
- Clausen, J. D., Christiansen, G., Holst, H. U. & Birkelund, S. Chlamydia trachomatis utilizes the host cell microtubule network during early events of infection. Mol. Microbiol.25, 441–449 (1997). - PubMed
-
- Grieshaber, S. S., Grieshaber, N. A. & Hackstadt, T. Chlamydia trachomatis uses host cell dynein to traffic to the microtubule-organizing center in a p50 dynamitin-independent process. J. Cell Sci.116, 3793–3802 (2003). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
