

Discovery of a single-subunit oligosaccharyltransferase that enables glycosylation of full-length IgG antibodies in bacteria
- PMID: 40610439
- PMCID: PMC12229607
- DOI: 10.1038/s41467-025-61440-7
Discovery of a single-subunit oligosaccharyltransferase that enables glycosylation of full-length IgG antibodies in bacteria
Abstract
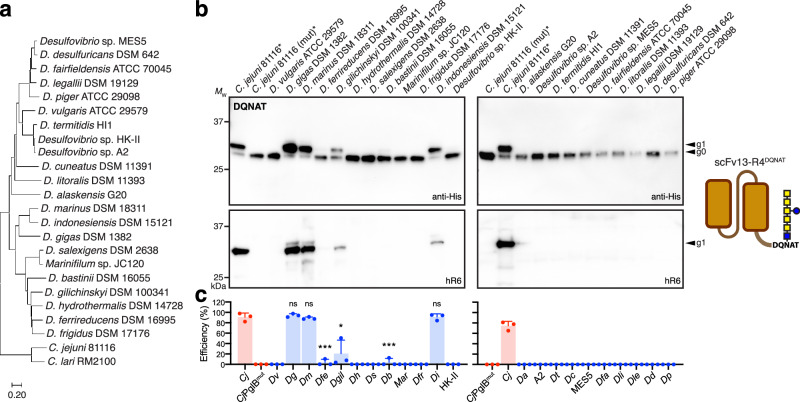
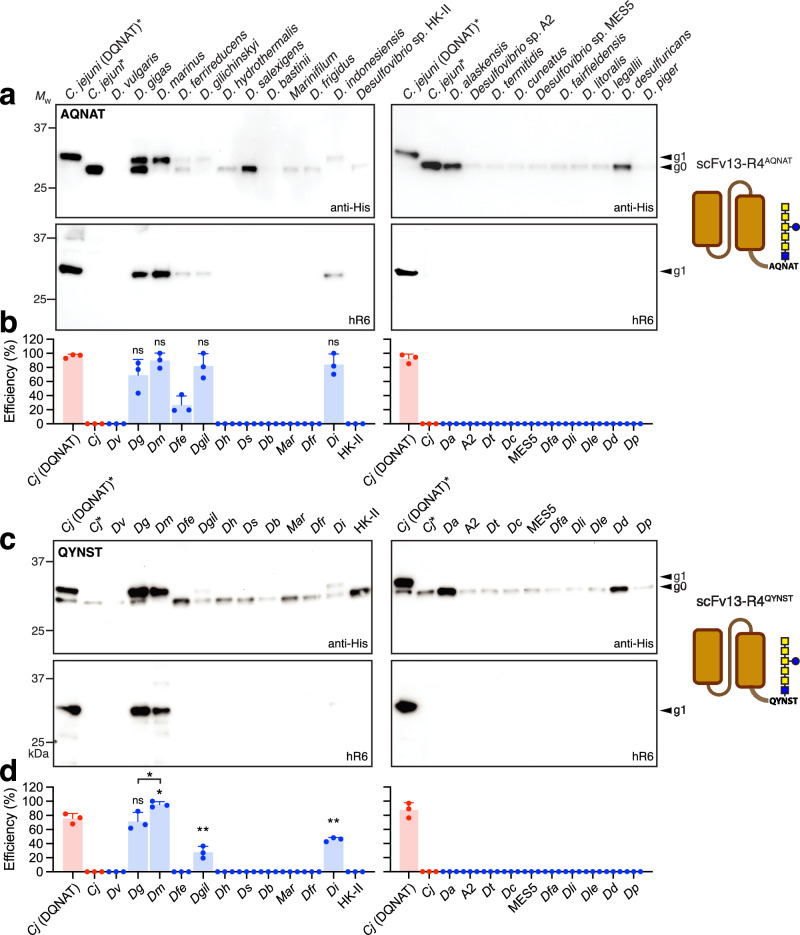
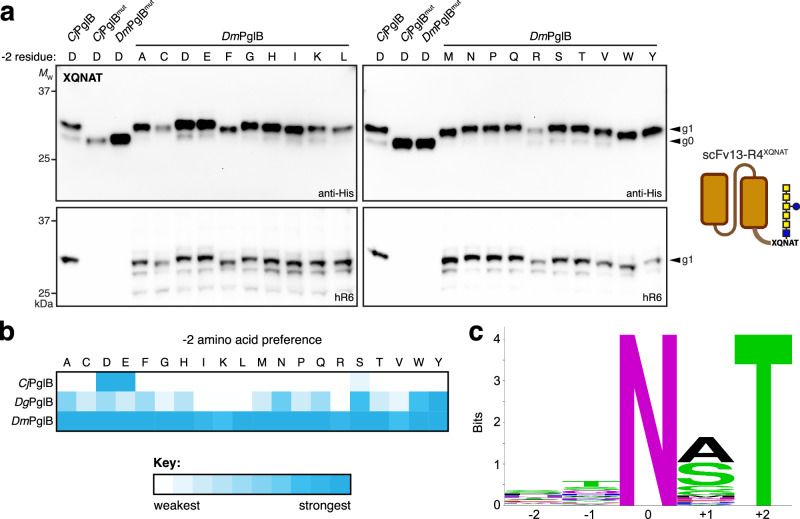
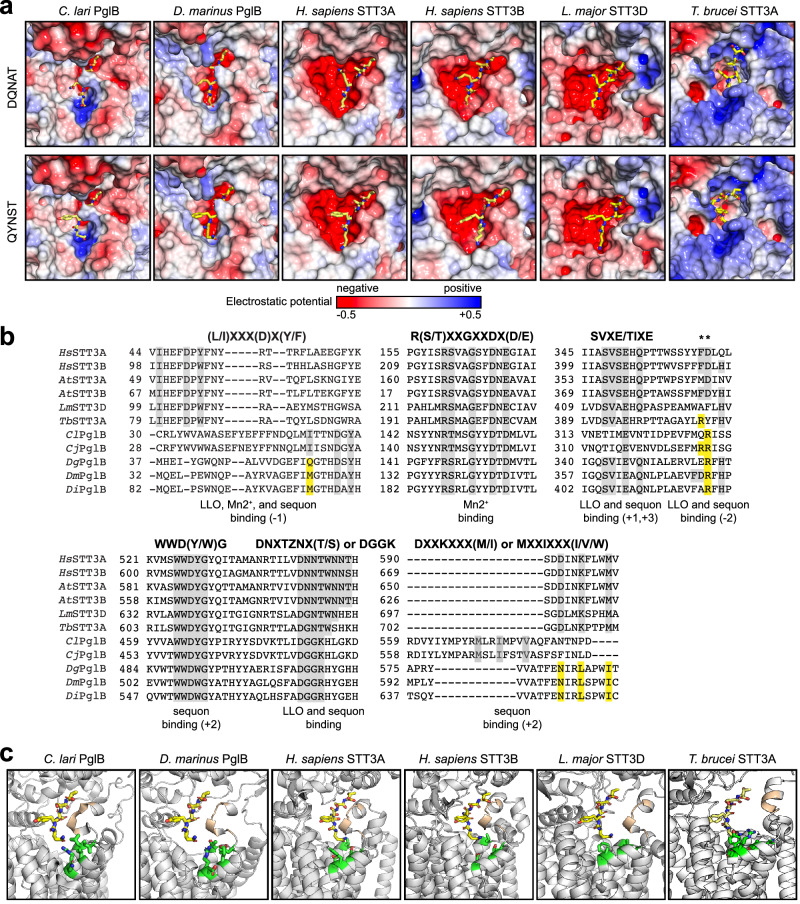
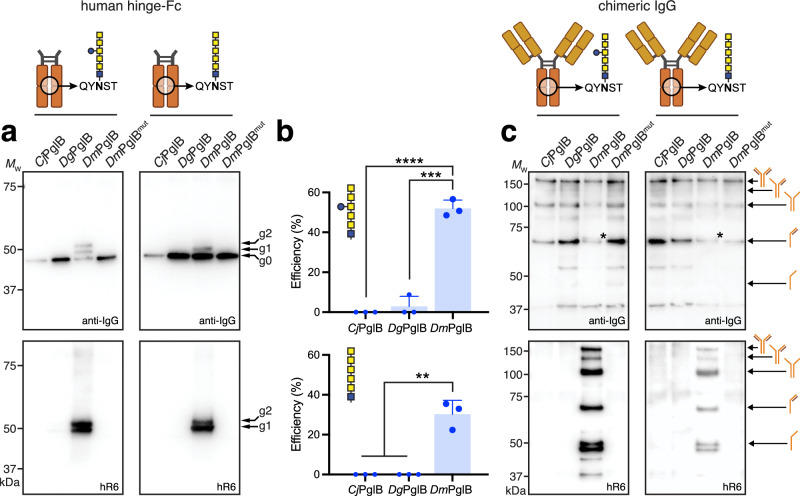
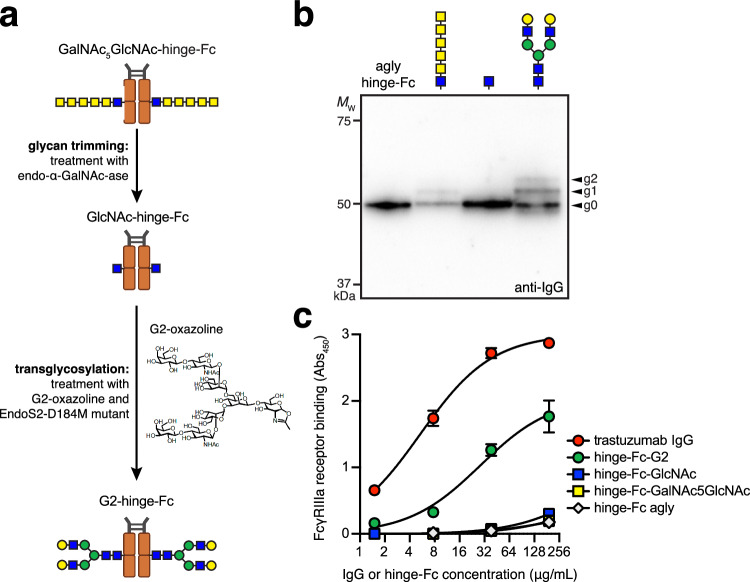
Human immunoglobulin G (IgG) antibodies are a major class of biotherapeutics and undergo N-linked glycosylation in their Fc domain, which is critical for immune functions and therapeutic activity. Hence, technologies for producing authentically glycosylated IgGs are in high demand. Previous attempts to engineer Escherichia coli for this purpose have met limited success due in part to the lack of oligosaccharyltransferase (OST) enzymes that can install N-glycans at the conserved N297 site in the Fc region. Here, we identify a single-subunit OST from Desulfovibrio marinus with relaxed substrate specificity that catalyzes glycosylation of native Fc acceptor sites. By chemoenzymatic remodeling the attached bacterial glycans to homogeneous, asialo complex-type G2 N-glycans, the E. coli-derived Fc binds human FcγRIIIa/CD16a, a key receptor for antibody-dependent cellular cytotoxicity (ADCC). Overall, the discovery of D. marinus OST provides previously unavailable biocatalytic capabilities and sets the stage for using E. coli to produce fully human antibodies.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: M.P.D. and M.C.J. have financial interests in Gauntlet, Inc. and Resilience, Inc. M.P.D. also has financial interests in June Bio APS, Glycobia Inc., UbiquiTx Inc., and Versatope Therapeutics Inc. M.P.D.’s and M.C.J.’s interests are reviewed and managed by Cornell University and Stanford University, respectively, in accordance with their conflict-of-interest policies. M.P.D. and M.C.J. declare no additional competing interests. All other authors declare no competing interests.
Figures
Update of
-
Discovery of a single-subunit oligosaccharyltransferase that enables glycosylation of full-length IgG antibodies in Escherichia coli.bioRxiv [Preprint]. 2024 Nov 10:2024.08.12.607630. doi: 10.1101/2024.08.12.607630. bioRxiv. 2024. Update in: Nat Commun. 2025 Jul 4;16(1):6152. doi: 10.1038/s41467-025-61440-7. PMID: 39574765 Free PMC article. Updated. Preprint.
Similar articles
-
Glycosylation of Structured Protein Domains in Cell-Free Reaction Environments.ACS Synth Biol. 2025 Jun 20;14(6):2354-2367. doi: 10.1021/acssynbio.5c00229. Epub 2025 May 28. ACS Synth Biol. 2025. PMID: 40437661 Free PMC article.
-
Discovery of a single-subunit oligosaccharyltransferase that enables glycosylation of full-length IgG antibodies in Escherichia coli.bioRxiv [Preprint]. 2024 Nov 10:2024.08.12.607630. doi: 10.1101/2024.08.12.607630. bioRxiv. 2024. Update in: Nat Commun. 2025 Jul 4;16(1):6152. doi: 10.1038/s41467-025-61440-7. PMID: 39574765 Free PMC article. Updated. Preprint.
-
Cell-based glycoengineering for production of homogeneous and specific glycoform-enriched antibodies with improved effector functions.Proc Natl Acad Sci U S A. 2025 Feb 25;122(8):e2423853122. doi: 10.1073/pnas.2423853122. Epub 2025 Feb 19. Proc Natl Acad Sci U S A. 2025. PMID: 39969996 Free PMC article.
-
Antibody tests for identification of current and past infection with SARS-CoV-2.Cochrane Database Syst Rev. 2022 Nov 17;11(11):CD013652. doi: 10.1002/14651858.CD013652.pub2. Cochrane Database Syst Rev. 2022. PMID: 36394900 Free PMC article.
-
Drugs for preventing postoperative nausea and vomiting in adults after general anaesthesia: a network meta-analysis.Cochrane Database Syst Rev. 2020 Oct 19;10(10):CD012859. doi: 10.1002/14651858.CD012859.pub2. Cochrane Database Syst Rev. 2020. PMID: 33075160 Free PMC article.
Cited by
-
Glycosylation of Structured Protein Domains in Cell-Free Reaction Environments.ACS Synth Biol. 2025 Jun 20;14(6):2354-2367. doi: 10.1021/acssynbio.5c00229. Epub 2025 May 28. ACS Synth Biol. 2025. PMID: 40437661 Free PMC article.
References
-
- Abu-Qarn, M., Eichler, J. & Sharon, N. Not just for Eukarya anymore: protein glycosylation in bacteria and archaea. Curr. Opin. Struct. Biol.18, 544–550 (2008). - PubMed
-
- Apweiler, R., Hermjakob, H. & Sharon, N. On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim. Biophys. Acta1473, 4–8 (1999). - PubMed
-
- Stanley, P., Moremen, K. W., Lewis, N. E., Taniguchi, N. & Aebi, M. In Essentials of Glycobiology, Edn. 4th. (eds. Varki, A. et al.) 103−116 (Cold Spring Harbor (NY), 2022).
-
- Seeberger, P.H., Freedberg, D.I. & Cummings, R.D. In Essentials of Glycobiology, 4th Edn. (eds. Varki, A. et al.) 771−784 (Cold Spring Harbor (NY), 2022).
MeSH terms
Substances
Grants and funding
- R01 GM127578/GM/NIGMS NIH HHS/United States
- R01GM127578, R01GM080374, R24GM137782/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- W911NF-23-2-0039/United States Department of Defense | Defense Advanced Research Projects Agency (DARPA)
- T32 GM138826/GM/NIGMS NIH HHS/United States
- R24 GM137782/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
