

Potential role of developmental experience in the emergence of the parvo-magno distinction
- PMID: 40610671
- PMCID: PMC12229566
- DOI: 10.1038/s42003-025-08382-4
Potential role of developmental experience in the emergence of the parvo-magno distinction
Abstract
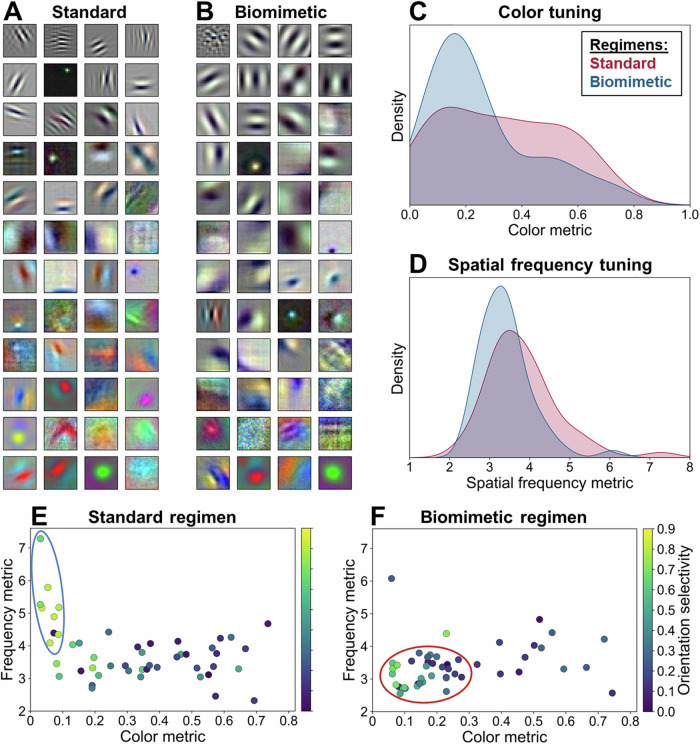
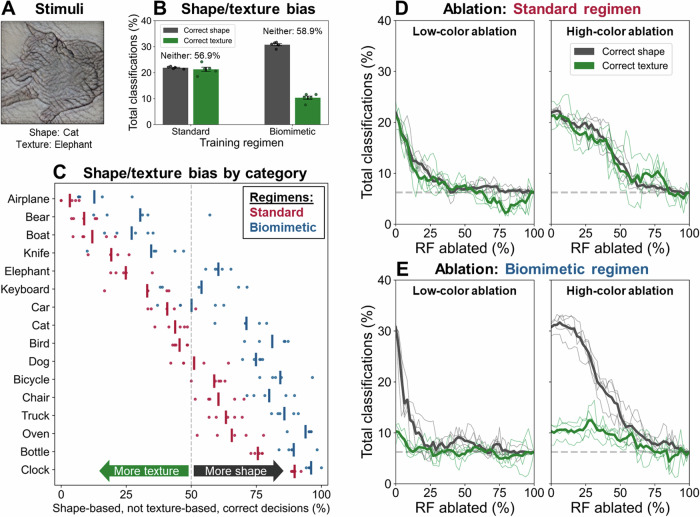
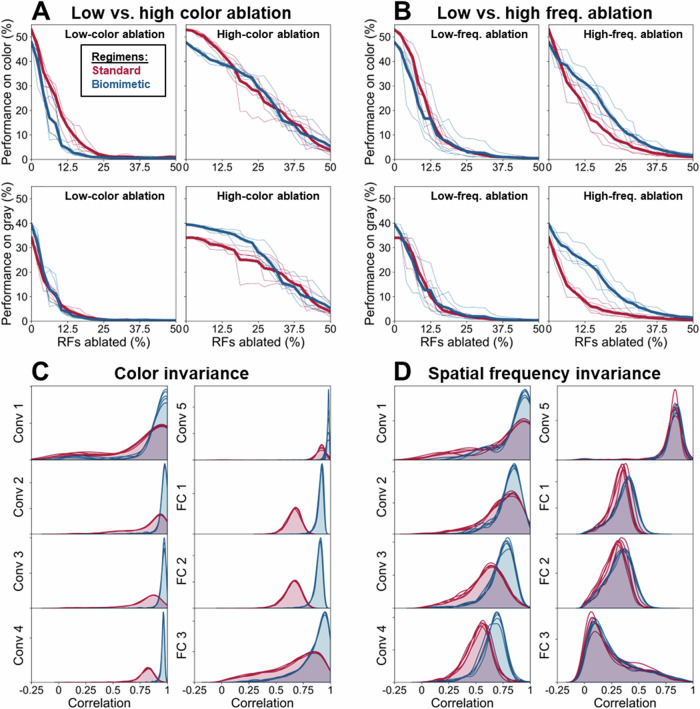
While the division of the early visual pathway into parvo- and magnocellular systems with distinct response properties has long been established as a prominent organizing principle in mammalian visual systems, the factors that lead to its emergence remain unclear. Here, we provide a potential account of this emergence based on early sensory development. Specifically, we propose that the temporal confluence in the developmental progression of spatial frequency and chromatic sensitivities may significantly shape corresponding neuronal response properties characteristic of this division. Receptive field analyses of deep networks trained on developmentally inspired 'biomimetic' protocols support this proposal in both the spatial and temporal domain. Further, biomimetic training induces a more human-like bias towards global shape processing, potentially driven by magnocellular units. These results have implications for the emergence of a key aspect of visual pathway organization and applied relevance for the design of training procedures for computational vision systems.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures

References
-
- Livingstone, M. & Hubel, D. Segregation of form, color, movement, and depth: anatomy, physiology, and perception. Science240, 740–749 (1988). - PubMed
-
- Shapley, R. Visual sensitivity and parallel retinocortical channels. Annu. Rev. Psychol.41, 635–658 (1990). - PubMed
-
- Shapley, R. Parallel retinocortical channels: X and Y and P and M. In Advances in psychology(Vol. 86, pp. 3-36). North-Holland (1992).
-
- Wiesel, T. N. & Hubel, D. H. Spatial and chromatic interactions in the lateral geniculate body of the rhesus monkey. J. Neurophysiol.29, 1115–1156 (1966). - PubMed
-
- Schiller, P. H. & Malpeli, J. G. Functional specificity of lateral geniculate nucleus laminae of the rhesus monkey. J. Neurophysiol.41, 788–797 (1978). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
