

Immunocompetent cell targeting by food-additive titanium dioxide
- PMID: 40615368
- PMCID: PMC12227633
- DOI: 10.1038/s41467-025-60248-9
Immunocompetent cell targeting by food-additive titanium dioxide
Abstract
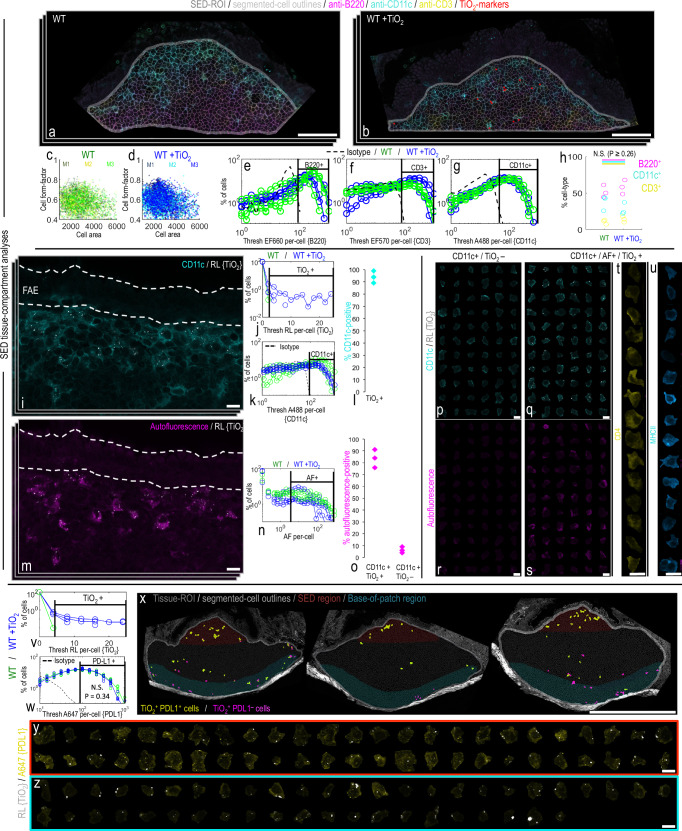
Food-grade titanium dioxide (fgTiO2) is a bio-persistent particle under intense regulatory scrutiny. Yet paradoxically, the only known cell reservoirs for fgTiO2 are graveyard intestinal pigment cells which are metabolically and immunologically quiescent. Here we identify immunocompetent cell targets of fgTiO2 in humans, most notably in the subepithelial dome region of intestinal Peyer's patches. Using multimodal microscopies with single-particle detection and per-cell / vesicle image analysis we achieve correlative dosimetry, quantitatively recapitulating human cellular exposures in the ileum of mice fed a fgTiO2-containing diet. Epithelial microfold cells selectively funnel fgTiO2 into LysoMac and LysoDC cells with ensuing accumulation. Notwithstanding, proximity extension analyses for 92 protein targets reveal no measureable perturbation of cell signalling pathways. When chased with oral ΔaroA-Salmonella, pro-inflammatory signalling is confirmed, but no augmentation by fgTiO2 is revealed despite marked same-cell loading. Interestingly, Salmonella causes the fgTiO2-recipient cells to migrate within the patch and, sporadically, to be identified in the lamina propria, thereby fully recreating the intestinal tissue distribution of fgTiO2 in humans. Immunocompetent cells that accumulate fgTiO2 in vivo are now identified and we demonstrate a mouse model that finally enables human-relevant risk assessments of ingested, bio-persistent (nano)particles.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: J.W. states that after completing the presented work as University of Cambridge Herchel-Smith Fellow, he subsequently undertook employment with the pharmaceutical company, GlaxoSmithKline. A.D., J.R., M.M., S.R., H.D.S., R.E.H., A.F., A.B.D.S., C.A.P.B., S.M., Å.V.K., J.D.S., N.C.R., D.O., R.J., P.M., A.P.B., P.R. and J.J.P. declare no competing interests.
Figures






References
-
- Ashwood, P., Thompson, R. P. & Powell, J. J. Fine particles that adsorb lipopolysaccharide via bridging calcium cations may mimic bacterial pathogenicity towards cells. Exp. Biol. Med.232, 107–117 (2007). - PubMed
-
- Grand View Research. Titanium dioxide market size, share and trends analysis report by grade (anatase, rutile), by production process (sulfate, chloride), by application (paints and coatings, plastics), by region, and segment forecasts, 2021−2028. https://www.grandviewresearch.com/industry-analysis/titanium-dioxide-ind... (2025).
-
- European Medicines Agency. Final feedback from European Medicines Agency (EMA) to the EU Commission request to evaluate the impact of the removal of titanium dioxide from the list of authorised food additives on medicinal products. https://www.ema.europa.eu/en/documents/report/final-feedback-european-me... (2025).
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
