

A plant bunyaviral protein disrupts SERRATE phase separation to modulate microRNA biogenesis during viral pathogenesis
- PMID: 40615422
- PMCID: PMC12227672
- DOI: 10.1038/s41467-025-61528-0
A plant bunyaviral protein disrupts SERRATE phase separation to modulate microRNA biogenesis during viral pathogenesis
Abstract
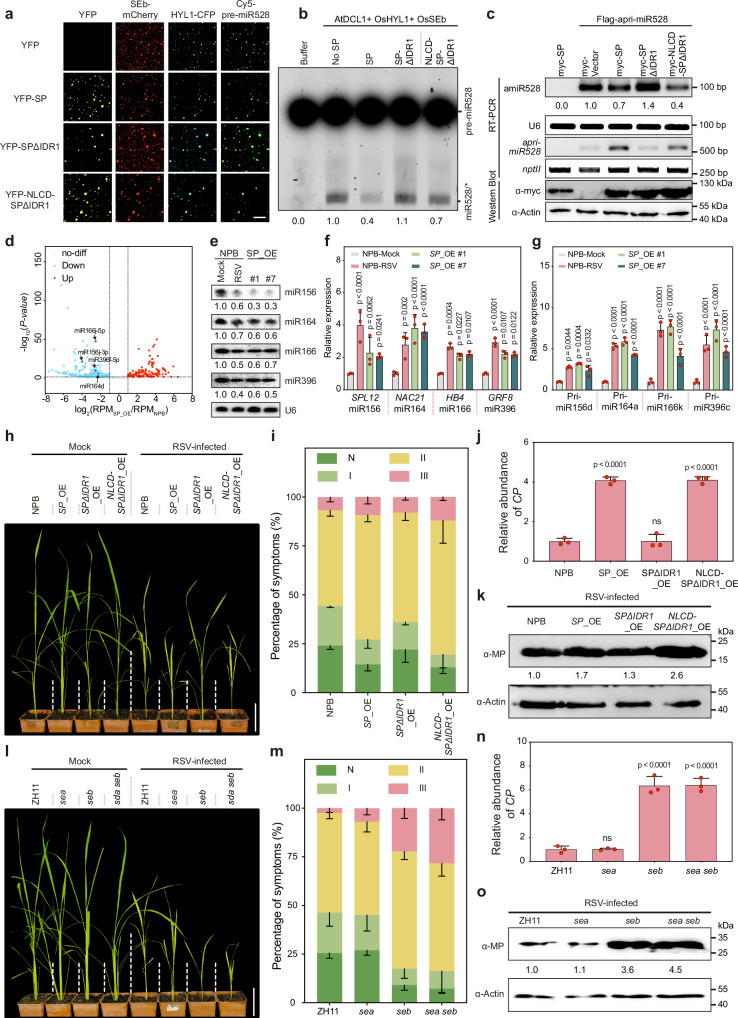
Liquid-liquid phase separation (LLPS) regulates diverse biological functions by mediating the assembly of biomolecular condensates. However, it remains unclear how host LLPS is targeted by viruses during infection. Here we show that a plant bunyaviral protein, the disease-specific protein (SP) encoded by rice stripe virus (RSV), possesses phase separation potential through its N-terminal intrinsically disordered region 1 (IDR1). In vivo, however, SP does not form phase-separated biomolecular condensates independently but utilizes its phase separation properties to interfere with the phase separation of the SERRATE protein (SE), a key component of Dicing bodies essential for microRNA processing. By disrupting SE phase separation, SP inhibits D-body assembly and miRNA biogenesis. Our study demonstrates that a viral protein can modulate host microRNA processing by targeting LLPS, revealing a previously uncharacterized mechanism involved in viral infection strategies and miRNA biogenesis regulation in plants.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures





Similar articles
-
Viral proteins suppress rice defenses by boosting OsTSN1 RNA decay via phase separation and multimerization.Nat Commun. 2025 Aug 12;16(1):7481. doi: 10.1038/s41467-025-62395-5. Nat Commun. 2025. PMID: 40796552 Free PMC article.
-
The H1/H5 domain contributes to OsTRBF2 phase separation and gene repression during rice development.Plant Cell. 2024 Sep 3;36(9):3787-3808. doi: 10.1093/plcell/koae199. Plant Cell. 2024. PMID: 38976557 Free PMC article.
-
Engineered Biomolecular Condensates Limit Tobacco Mosaic Virus Accumulation and Symptom Development.Mol Plant Pathol. 2025 Jun;26(6):e70113. doi: 10.1111/mpp.70113. Mol Plant Pathol. 2025. PMID: 40544335 Free PMC article.
-
Small Molecules as Regulators of Liquid-Liquid Phase Separation: Mechanisms and Strategies for New Drug Discovery.FASEB J. 2025 Jul 15;39(13):e70773. doi: 10.1096/fj.202501476R. FASEB J. 2025. PMID: 40616404 Review.
-
A story of two kingdoms: unravelling the intricacies of protein phase separation in plants and animals.Crit Rev Biotechnol. 2025 Aug;45(5):1019-1039. doi: 10.1080/07388551.2024.2425989. Epub 2024 Nov 26. Crit Rev Biotechnol. 2025. PMID: 39592156 Review.
References
MeSH terms
Substances
Grants and funding
- U1905203/National Natural Science Foundation of China (National Science Foundation of China)
- 32300149/National Natural Science Foundation of China (National Science Foundation of China)
- 32072381/National Natural Science Foundation of China (National Science Foundation of China)
- 2024J01378/Natural Science Foundation of Fujian Province (Fujian Provincial Natural Science Foundation)
LinkOut - more resources
Full Text Sources
