

Enhancer adoption by an LTR retrotransposon generates viral-like particles, causing developmental limb phenotypes
- PMID: 40634706
- PMCID: PMC12283350
- DOI: 10.1038/s41588-025-02248-5
Enhancer adoption by an LTR retrotransposon generates viral-like particles, causing developmental limb phenotypes
Abstract
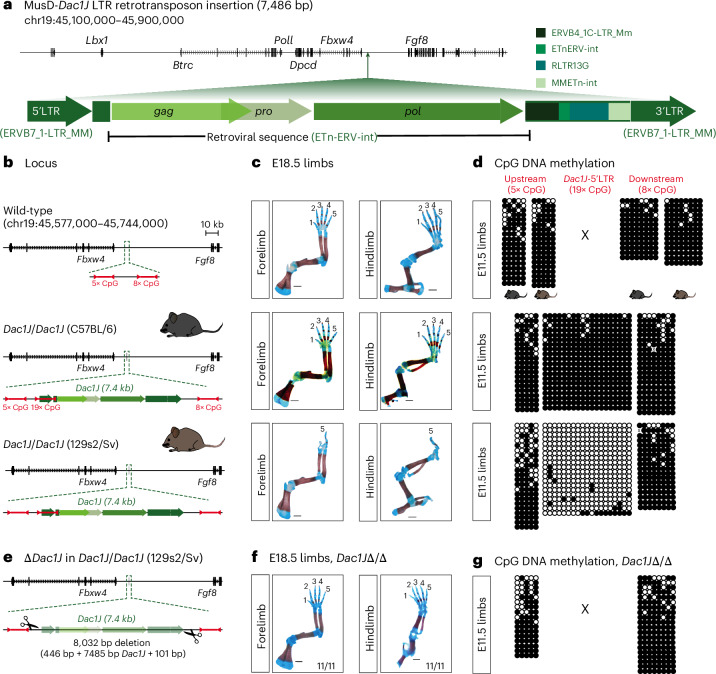
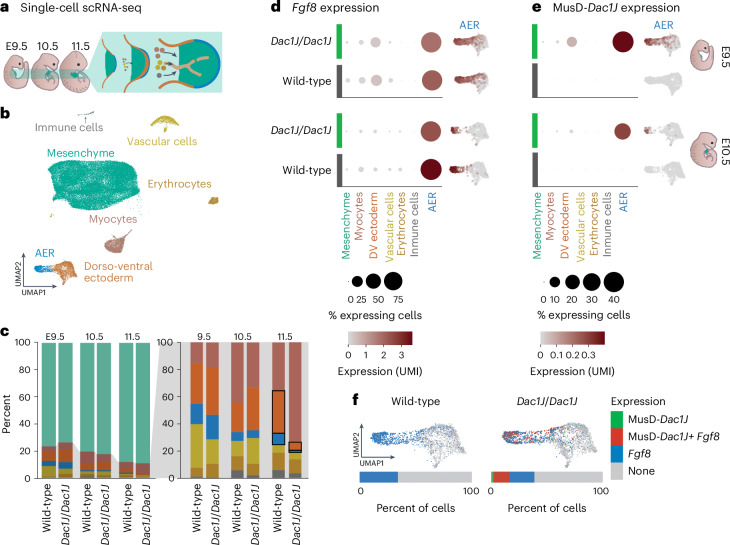
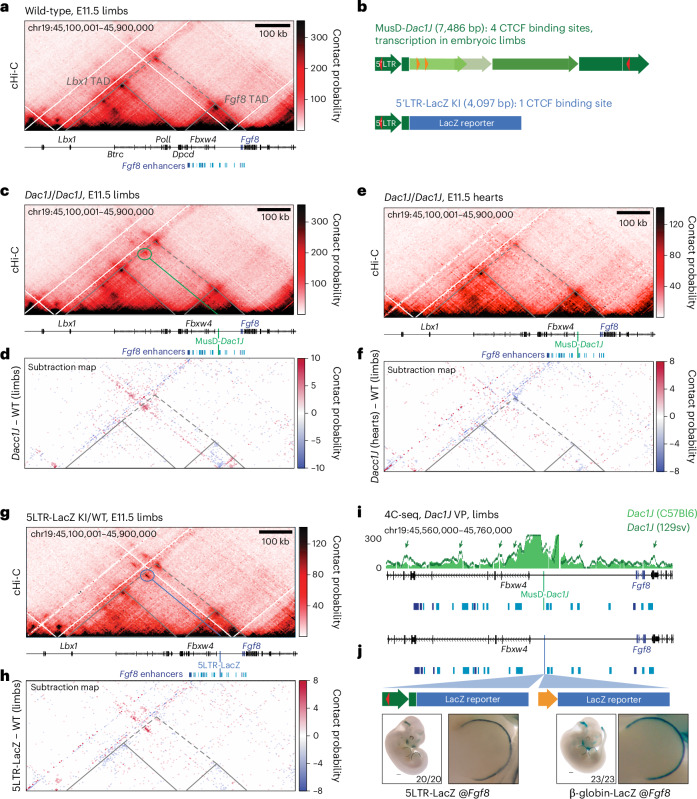
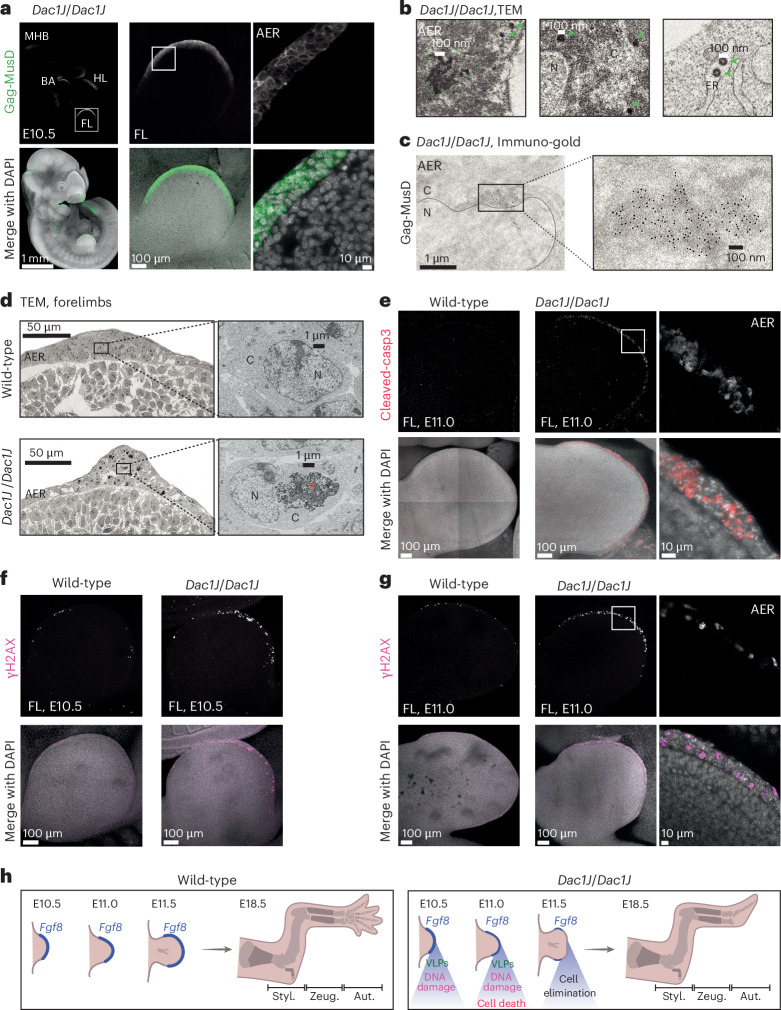
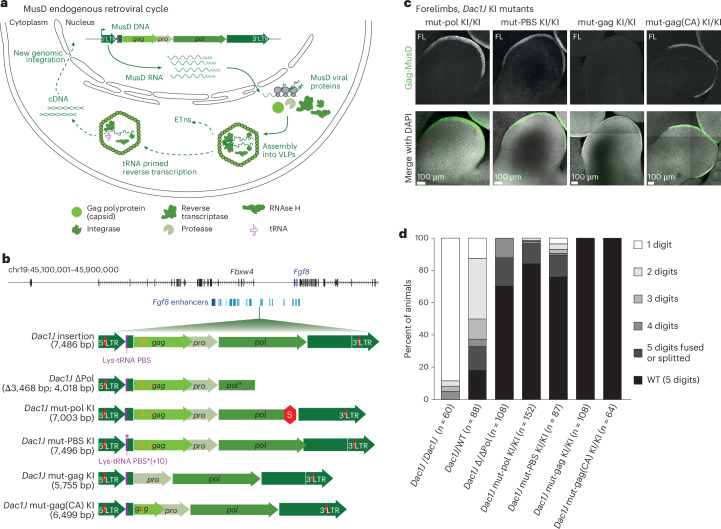
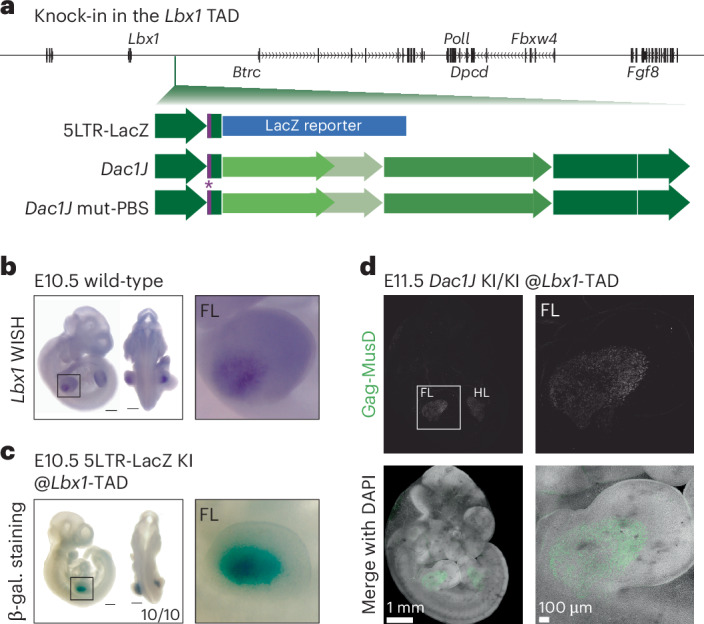
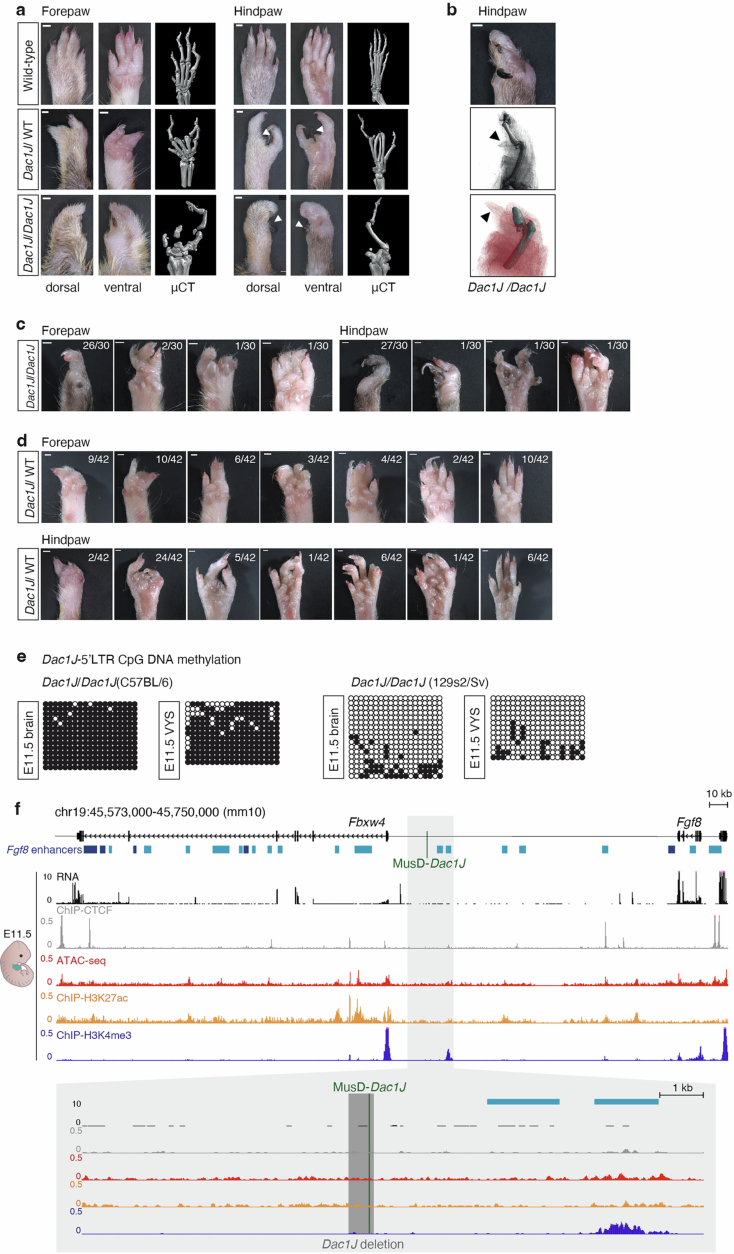
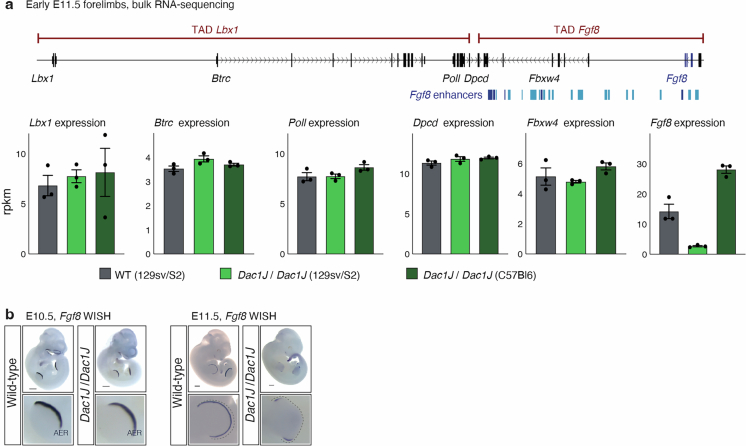
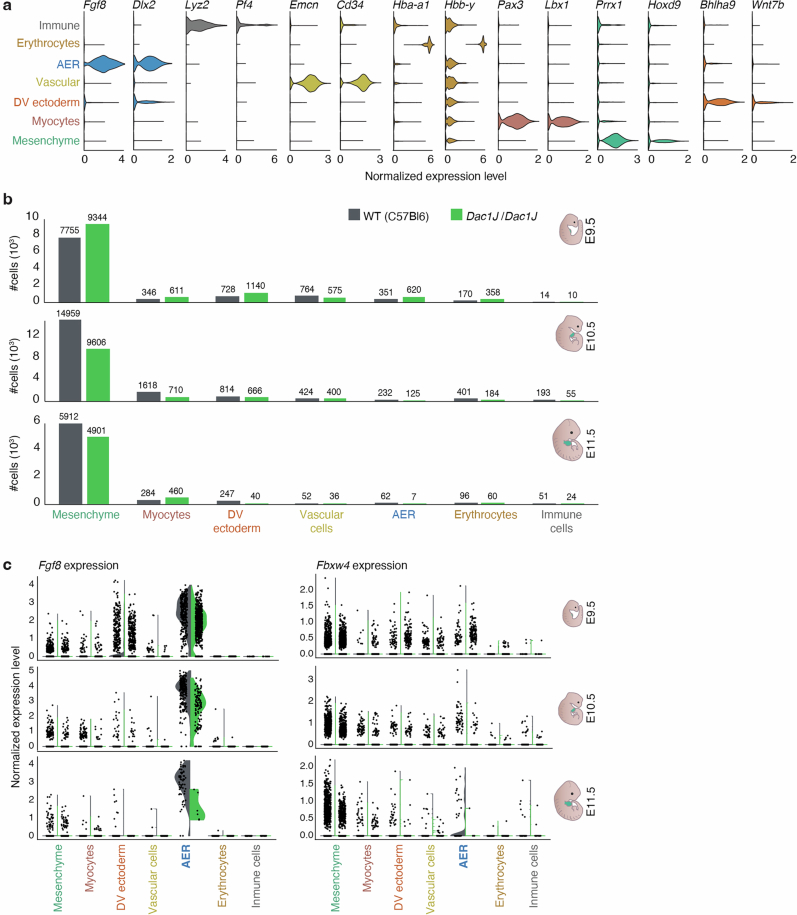
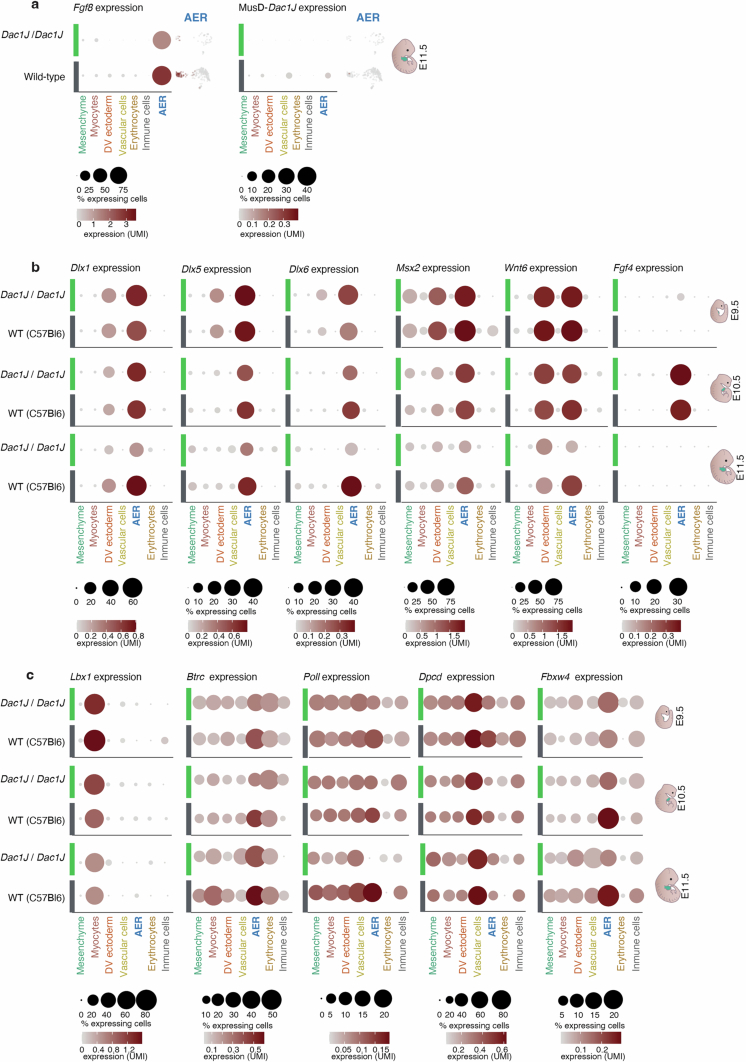
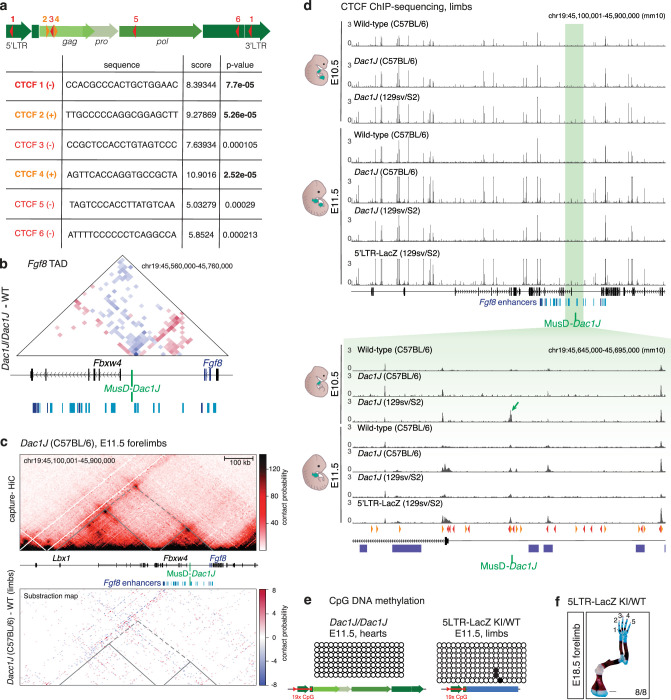
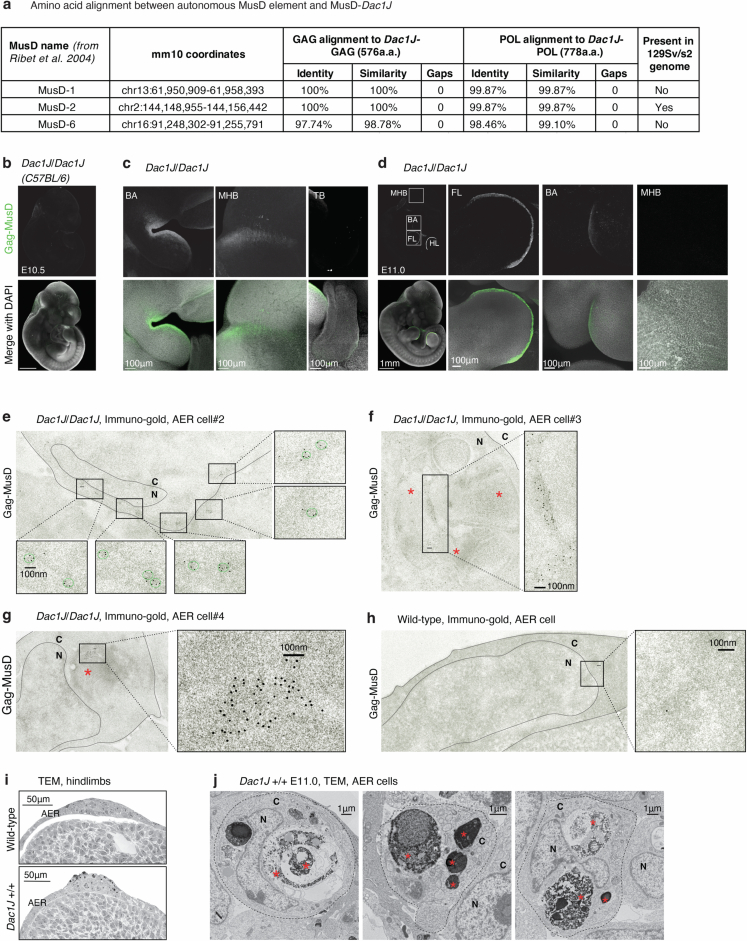
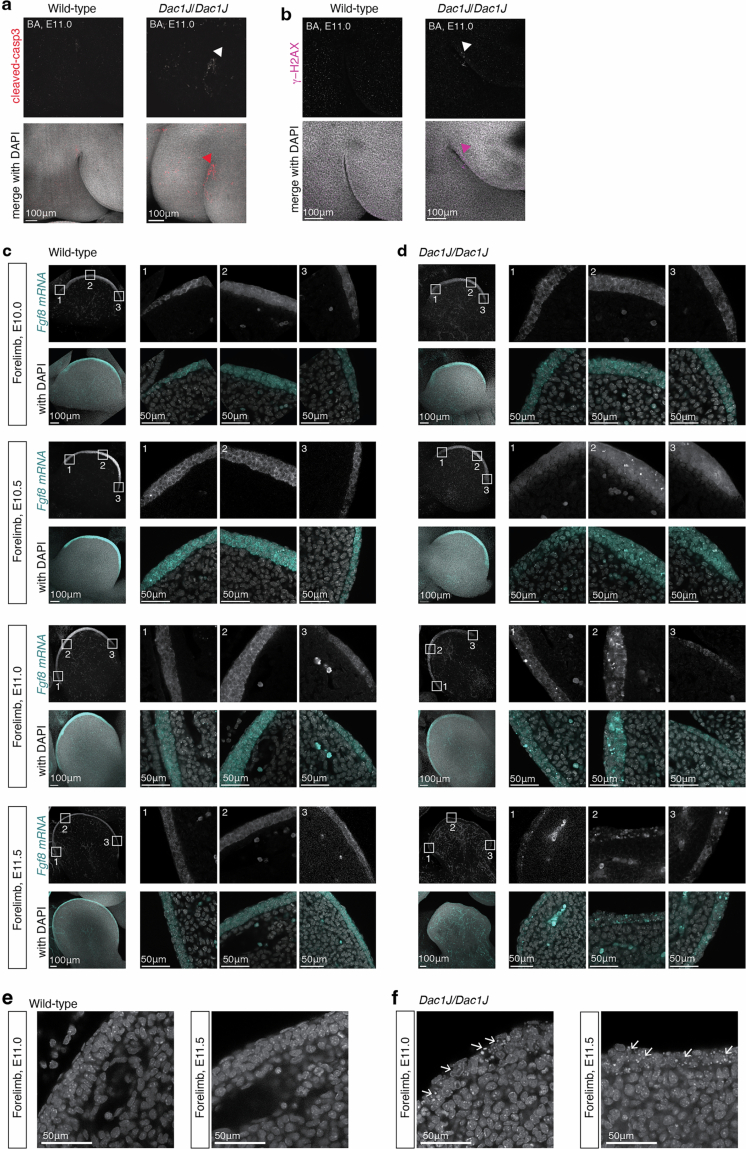
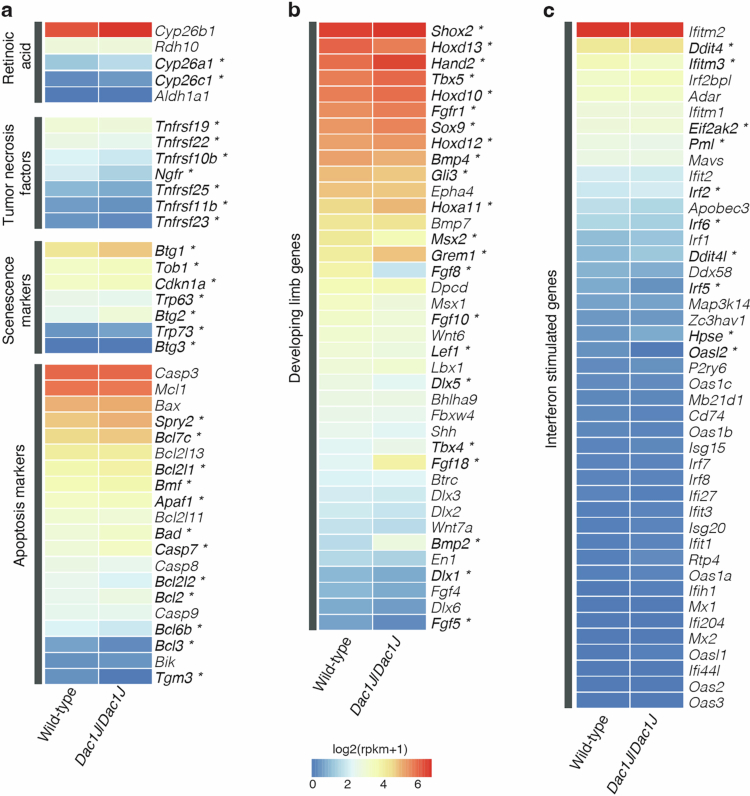
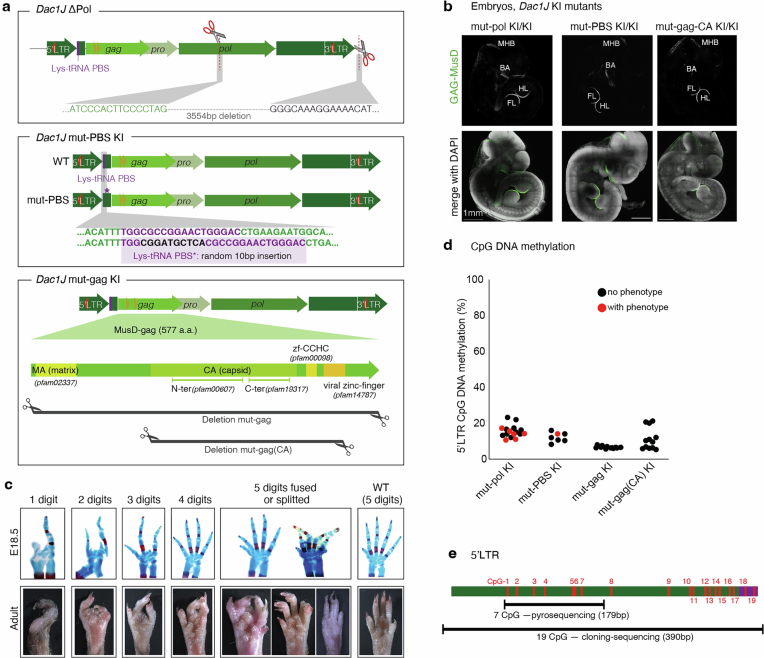
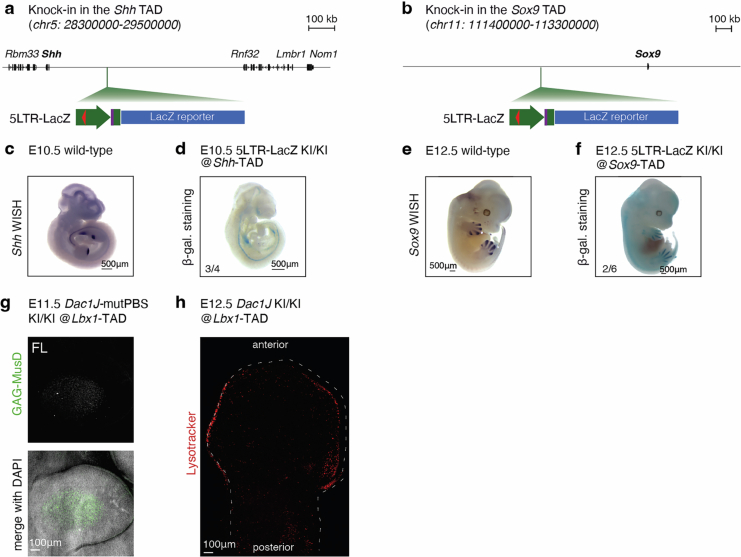
Transposable elements (TEs) are scattered across mammalian genomes. Silencing of TEs prevents harmful effects caused by either global activation leading to genome instability or insertional mutations disturbing gene transcription. However, whether the activation of a TE can cause disease without directly affecting gene expression is largely unknown. Here we show that a TE insertion can adopt nearby regulatory activity, resulting in the production of cell-type-specific viral-like particles (VLPs) that affect embryo formation. Failure to silence an LTR retrotransposon inserted upstream of the Fgf8 gene results in their co-expression during mouse development. VLP assembly in the Fgf8-expressing cells of the developing limb triggers apoptotic cell death, resulting in a limb malformation resembling human ectrodactyly. The phenotype can be rescued by mutating the retrotransposon coding sequence, thus preventing its full endogenous retroviral cycle. Our findings illustrate that TE insertions can be incorporated into the local genomic regulatory landscape and that VLP production in post-implantation embryos can cause developmental defects.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures
Similar articles
-
Repeat competition and ecological shifts drive the evolution of the mobilome in Rhynchospora Vahl (Cyperaceae), the holocentric beaksedges.Ann Bot. 2025 May 9;135(5):909-924. doi: 10.1093/aob/mcae220. Ann Bot. 2025. PMID: 39693387
-
Genetically regulated epigenetic transcriptional activation of retrotransposon insertion confers mouse dactylaplasia phenotype.Proc Natl Acad Sci U S A. 2007 Nov 27;104(48):19034-9. doi: 10.1073/pnas.0705483104. Epub 2007 Nov 5. Proc Natl Acad Sci U S A. 2007. PMID: 17984064 Free PMC article.
-
Comparative Analysis of Transposable Element Evolution in Crustaceans.Genome Biol Evol. 2025 Jul 3;17(7):evaf115. doi: 10.1093/gbe/evaf115. Genome Biol Evol. 2025. PMID: 40462399 Free PMC article.
-
Gonadotropin-releasing hormone (GnRH) analogues for premenstrual syndrome (PMS).Cochrane Database Syst Rev. 2025 Jun 10;6(6):CD011330. doi: 10.1002/14651858.CD011330.pub2. Cochrane Database Syst Rev. 2025. PMID: 40492482 Review.
-
Management of urinary stones by experts in stone disease (ESD 2025).Arch Ital Urol Androl. 2025 Jun 30;97(2):14085. doi: 10.4081/aiua.2025.14085. Epub 2025 Jun 30. Arch Ital Urol Androl. 2025. PMID: 40583613 Review.
Cited by
-
A retrotransposon caught red-handed in a curious case of missing digits.Nat Genet. 2025 Jul;57(7):1571-1573. doi: 10.1038/s41588-025-02147-9. Nat Genet. 2025. PMID: 40634705 No abstract available.
References
-
- Payer, L. M. & Burns, K. H. Transposable elements in human genetic disease. Nat. Rev. Genet.20, 760–772 (2019). - PubMed
-
- Deniz, Ö., Frost, J. M. & Branco, M. R. Regulation of transposable elements by DNA modifications. Nat. Rev. Genet.20, 417–431 (2019). - PubMed
-
- Fadloun, A. et al. Chromatin signatures and retrotransposon profiling in mouse embryos reveal regulation of LINE-1 by RNA. Nat. Struct. Mol. Biol.20, 332–338 (2013). - PubMed
-
- Jachowicz, J. W. et al. LINE-1 activation after fertilization regulates global chromatin accessibility in the early mouse embryo. Nat. Genet.49, 1502–1510 (2017). - PubMed
MeSH terms
Substances
Grants and funding
- MU 880/16-1/Deutsche Forschungsgemeinschaft (German Research Foundation)
- LT000465/2019-L/Human Frontier Science Program (HFSP)
- PP00P3_176802/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
- PP00P3_210995-6/National Centre of Competence in Research Robotics (Swiss National Center of Competence in Research "Robotics")
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
