

HP1 loses its chromatin clustering and phase separation function across evolution
- PMID: 40640210
- PMCID: PMC12246093
- DOI: 10.1038/s41467-025-61749-3
HP1 loses its chromatin clustering and phase separation function across evolution
Abstract
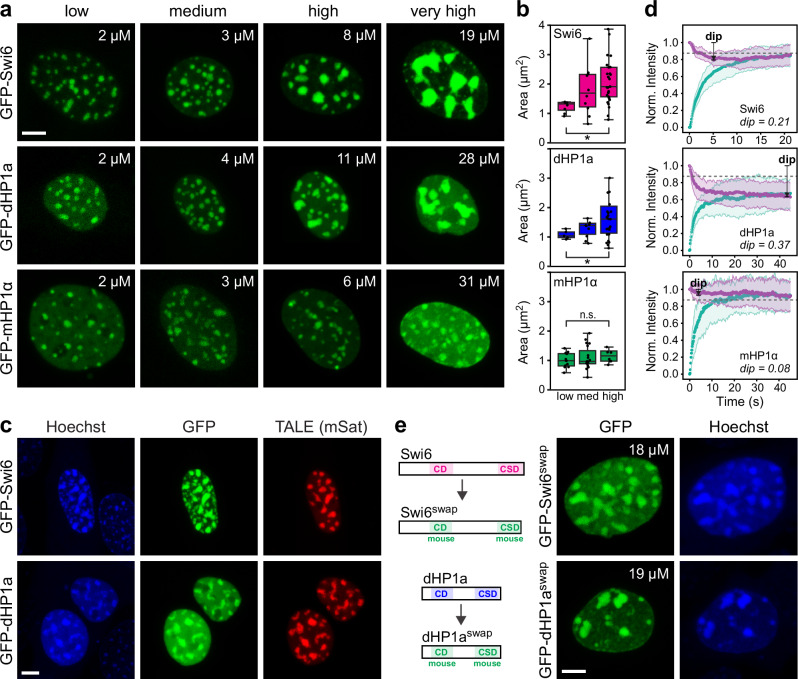
Heterochromatin protein 1 (HP1) is a multifunctional chromatin-associated protein conserved from fission yeast to mammals. HP1 has been suggested to drive heterochromatin formation via phase separation. However, there is seemingly conflicting evidence about HP1 phase-separating in different systems or not. Here, we assess the phase separation behavior of HP1 from fission yeast, fruit fly and mouse in vitro and in mammalian cells side-by-side. We find that HP1 from fission yeast and fly can undergo liquid-liquid phase separation and induce heterochromatin coalescence in mouse cells, in stark contrast to HP1 from mouse. Induced heterochromatin coalescence has only mild effects on gene expression. We link the decreasing phase separation propensity of HP1 homologs to their decreasing intrinsic disorder and their increasing sensitivity to HP1 paralogs antagonizing phase separation. Our work elucidates the relationship between phase separation, nuclear organization and gene expression, and highlights the evolutionary dimension of protein phase separation control.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures





References
-
- Lorentz, A., Ostermann, K., Fleck, O. & Schmidt, H. Switching gene swi6, involved in repression of silent mating-type loci in fission yeast, encodes a homologue of chromatin-associated proteins from Drosophila and mammals. Gene143, 139–143 (1994). - PubMed
-
- Jacobs, S. A. & Khorasanizadeh, S. Structure of HP1 chromodomain bound to a lysine 9-methylated histone H3 tail. Science295, 2080–2083 (2002). - PubMed
-
- Nielsen, P. R. et al. Structure of the HP1 chromodomain bound to histone H3 methylated at lysine 9. Nature416, 103–107 (2002). - PubMed
MeSH terms
Substances
Grants and funding
- 804023/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- 101170239/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
