

Complex I inhibition combined with TLR activation in the breast tumor microenvironment educates cytotoxic neutrophils
- PMID: 40644540
- PMCID: PMC12248295
- DOI: 10.1126/sciadv.adu5915
Complex I inhibition combined with TLR activation in the breast tumor microenvironment educates cytotoxic neutrophils
Abstract
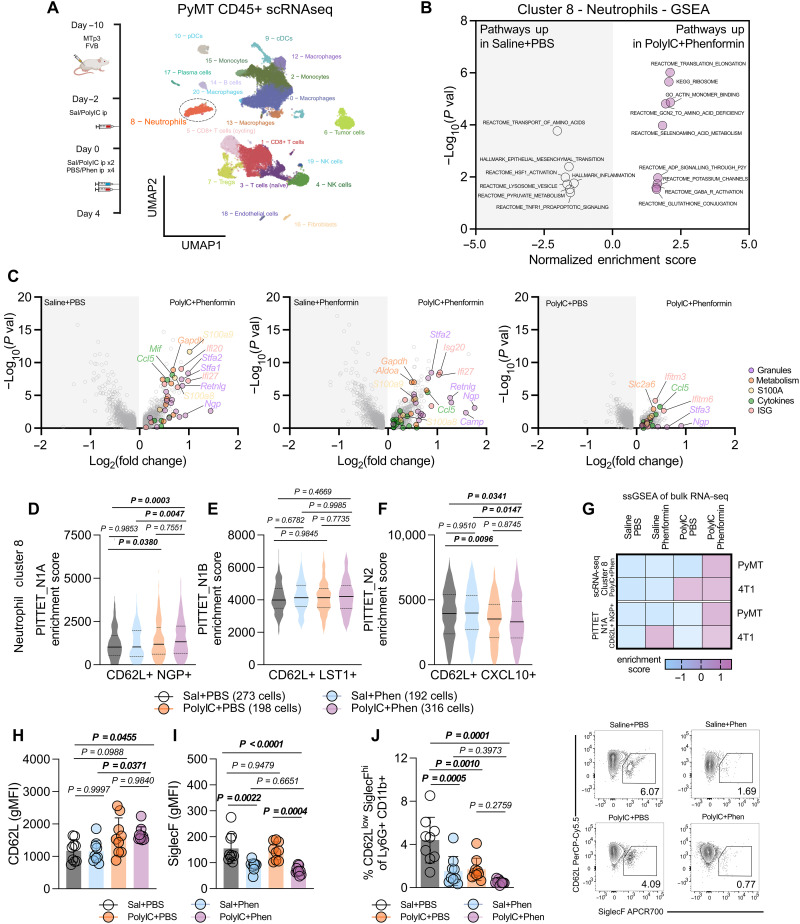
Although effective for immunologically hot tumors, immune checkpoint inhibitors minimally affect tumors that are not T cell inflamed, including breast cancer. An alternate strategy to combat immune cold breast tumors may be to reeducate innate immunity. This study identifies strategies to skew neutrophils to acquire tumoricidal properties. Systemic Toll-like receptor (TLR)-induced inflammation, concomitant with mitochondrial complex I inhibition in breast tumors, increases neutrophil cytotoxicity against breast cancer cells and independently of CD8+ T cell immunity. These therapy-entrained neutrophils enhance secretory granule production, increasing expression of the reduced form of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase machinery and inducing a respiratory burst. Moreover, systemic administration of TLR agonists elevates nuclear factor κB signaling in neutrophils to increase production of secretory granule and NADPH oxidase machinery components, whereas complex I inhibitors are required to potentiate oxidative damage. In summary, we describe a class of neutrophils, educated by the combined action of inflammatory mediators and metabolic inhibitors, having tumoricidal functions.
Figures








References
-
- Dagogo-Jack I., Shaw A. T., Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 15, 81–94 (2018). - PubMed
-
- de Visser K. E., Joyce J. A., The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell 41, 374–403 (2023). - PubMed
-
- Tong L., Chuang C. C., Wu S., Zuo L., Reactive oxygen species in redox cancer therapy. Cancer Lett. 367, 18–25 (2015). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
