

GPR101 loss promotes insulin resistance and diet-induced obesity risk
- PMID: 40655975
- PMCID: PMC12244024
- DOI: 10.1016/j.nsa.2023.101126
GPR101 loss promotes insulin resistance and diet-induced obesity risk
Abstract
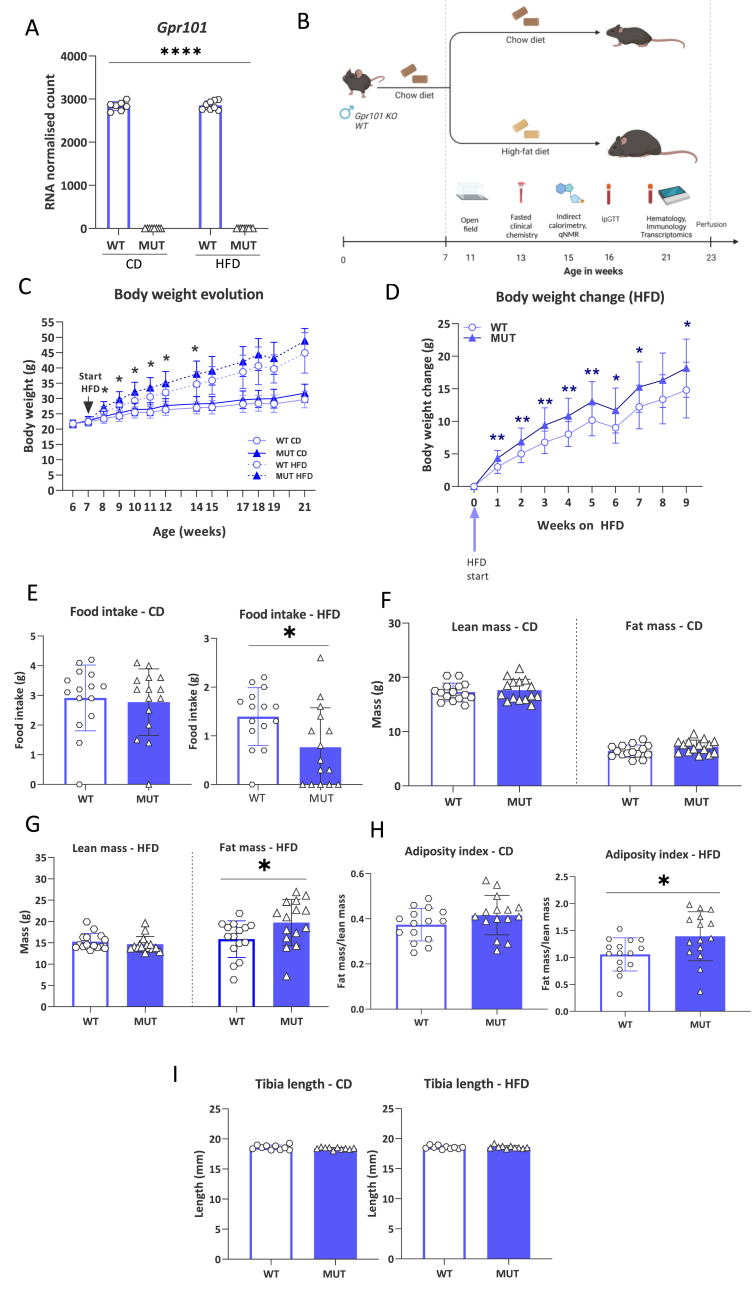
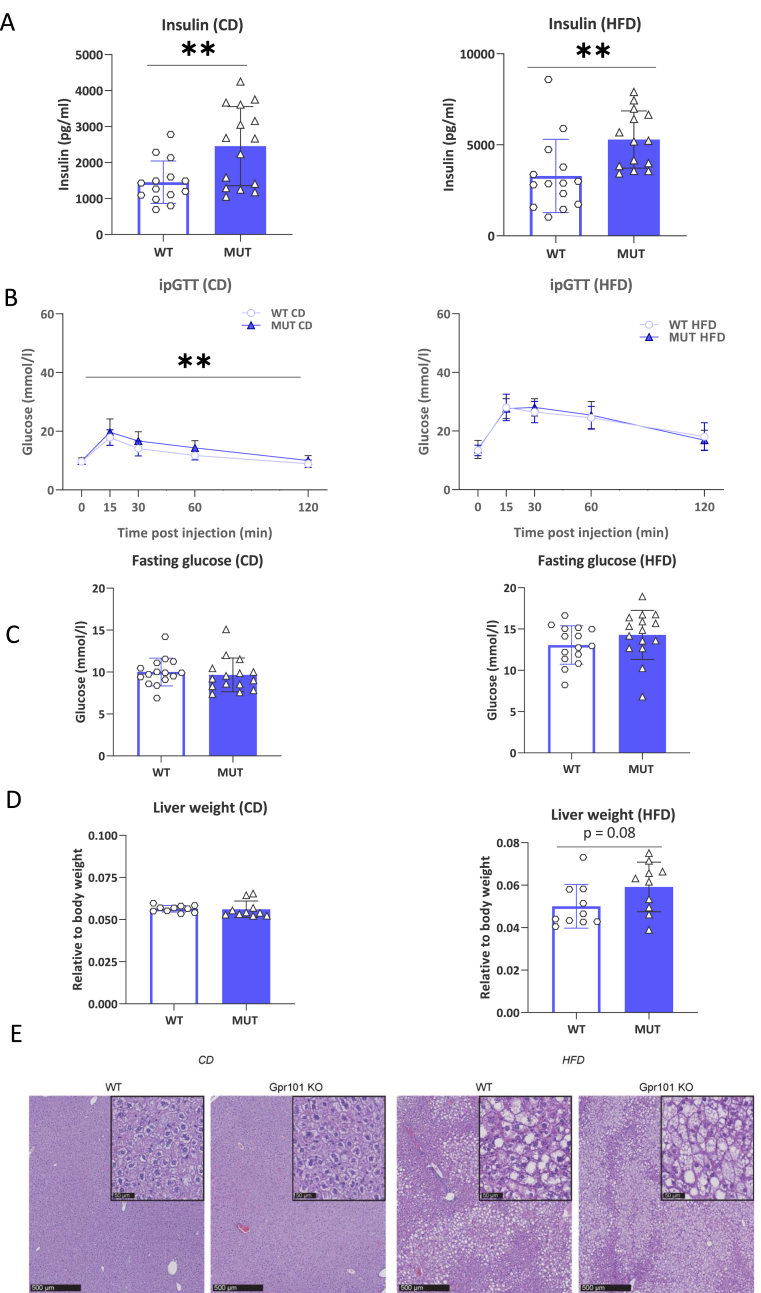
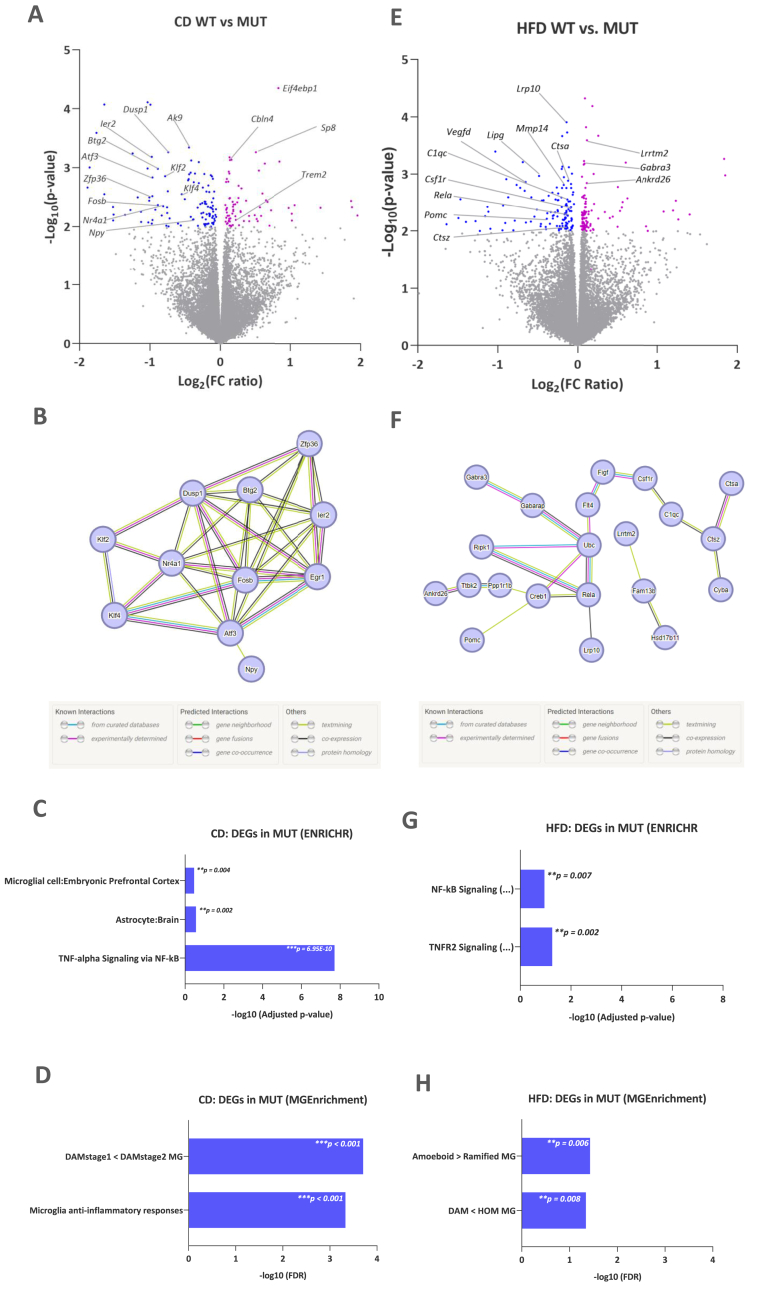
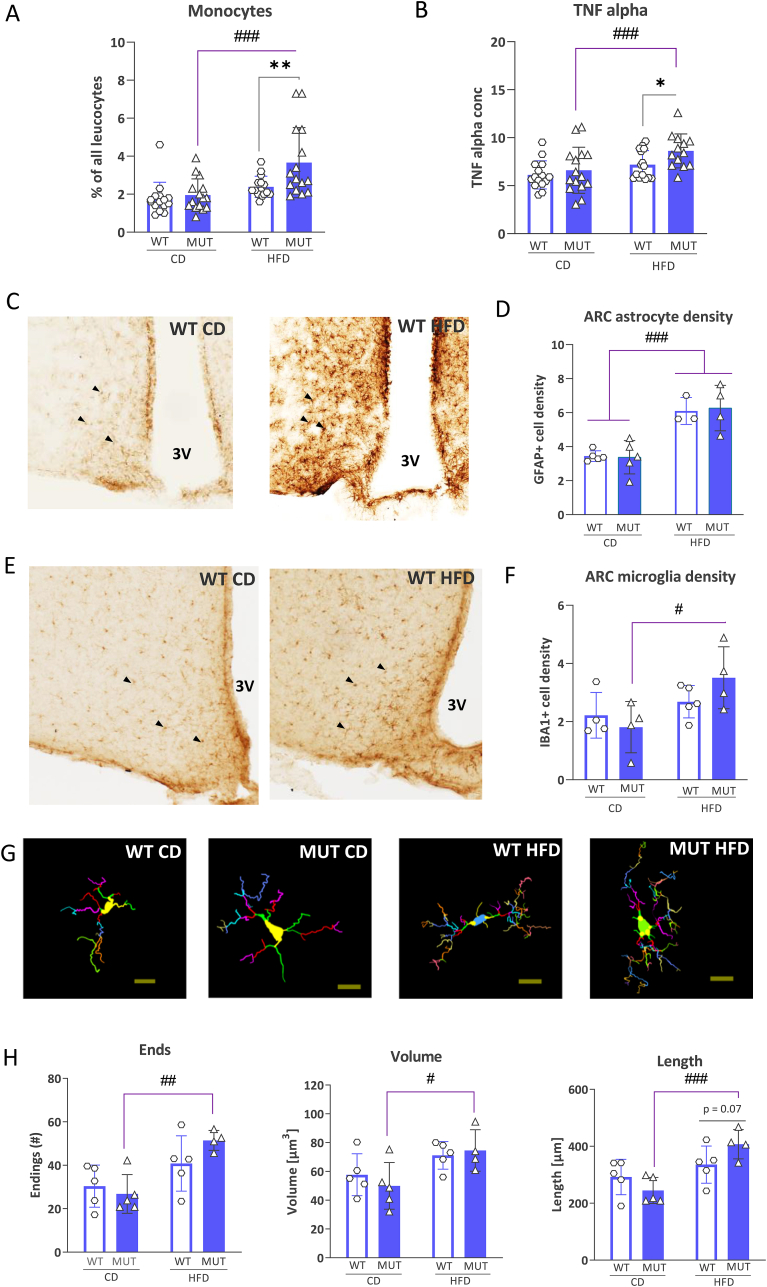
G-protein-coupled receptors (GPCRs) represent targets for improved low-side-effect therapies to tackle the evolving Western obesity epidemic. The orphan (o) GPCR GPR101 emerged as an attractive candidate in this regard. Expressed on cells in brain areas regulating energy homeostasis, including the hunger-suppressing proopiomelanocortin (POMC) + neurons, it is minimally expressed outside the brain. To understand the function of this receptor in vivo, we herein generated and comprehensively characterized a Gpr101 knockout mouse line, either under standard feeding conditions or with chronic high-fat diet (HFD) access (16 weeks). GPR101 loss accelerated the risk for diet-induced obesity (DIO), hyperinsulinemia and disrupted glucose homeostasis. Hypothalamic transcriptomic analysis revealed also decreased Pomc activation with HFD suggesting impaired hunger suppression. Moreover, on a standard diet, there was a molecular signature of downregulated tristetraprolin (TTP) interactome gene activation suggesting impaired inflammation resolution. On HFD, there was differential expression of genes involved in microglial phagocytosis and lipid metabolism. Morphometry revealed altered hypothalamic arcuate nucleus microglial morphology consistent with the transcriptomic profile. We discuss how the GPR101 specialized pro-resolving mediator (SPM) receptor capacity likely underlies the aberrant microglial function and contributes to DIO risk. Thus, this evidence shows that GPR101 is a potential therapeutic target for DIO through, among other factors, effects on hypothalamic inflammation resolution.
Keywords: Diet-induced obesity; GPR101; Hypothalamus; Inflammation; Insulin resistance.
© 2023 The Authors.
Conflict of interest statement
AB and TZ are employees of Boehringer Ingelheim Pharma GmbH & Co. KG. AB and TZ declare no competing financial interest in this work.The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Martin Hrabe de Angelis reports was provided by 10.13039/501100002347German Federal Ministry of Education and Research (Infrafrontier grant 01KX1012).
Figures
References
-
- Akiyama M., Okada Y., Kanai M., Takahashi A., Momozawa Y., Ikeda M., Iwata N., Ikegawa S., Hirata M., Matsuda K., Iwasaki M., Yamaji T., Sawada N., Hachiya T., Tanno K., Shimizu A., Hozawa A., Minegishi N., Tsugane S., Yamamoto M., Kubo M., Kamatani Y. Genome-wide association study identifies 112 new loci for body mass index in the Japanese population. Nat. Genet. 2017;49:1458–1467. - PubMed
-
- Bagnol D. US Patent Office (USPTO), U.S. patent No; 2010. Use of gpr101 Receptor in Methods to Identify Modulators of Hypothalamic Proopiomelanocortin (POMC)-derived Biologically Active Peptide Secretion Useful in the Treatment of Pomc-Derived Biologically Active Peptide-Related Disorders; pp. 1–80. 2010.
LinkOut - more resources
Full Text Sources
Miscellaneous